










Serviços Personalizados
artigo
 Português (pdf)
Português (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
Indicadores

Compartilhar

 Permalink
PermalinkPsicologia USP
versão On-line ISSN 1678-5177
Psicol. USP v.6 n.1 São Paulo 1995
ARTIGOS ORIGINAIS
Divisão de trabalho e especialização em obreiras de Atta sexdens1
Division of labor and specialization in Atta sexdens workers
Maria Margarida P. Rodrigues
Departamento de Psicologia Social e do Desenvolvimento - Universidade Federal Espírito Santo
RESUMO
A divisão de trabalho em função da idade e do tamanho corporal já foi constatada em várias espécies de insetos sociais. No gênero Atta, Carvalho (1972) e Wilson (1980a, 1980b) encontraram evidências inequívocas da existência de polietismo - divisão de trabalho - entre as obreiras quanto à idade e tamanho corporal. Estudos com A. sexdens já demonstraram que obreiras pequenas - com menos de 3 mm - exibem preferencialmente atividades relativas ao fungo (Carvalho, 1972). A presente pesquisa teve por objetivo verificar como a remoção parcial das jardineiras afeta duas das atividades que ocorrem na esponja de fungo: jardinagem e cuidados com formas imaturas (ovos, pupas e larvas). Os dados obtidos mostraram que, sob condições que acarretam a diminuição do número de obreiras da subclasse jardineira, as obreiras das subclasses próximas aumentam sua atividade jardineira sem diminuir os cuidados com formas imaturas.
Descritores: Divisão de trabalho animal. Formigas. Diferenças de idade. Tamanho do corpo. Comportamento de forrageamento animal.
ABSTRACT
The division of labor according to age and body size has already been observed in several epecies of social insects. In the Atta genus, Carvalho (1972) and Wilson (1980a, 1980b) found unmistakable evidences of the existence of polyethism - division of labor - among workers according to age and body size. Studies with Atta sexdens have proved that small workers - less than 3 mm long - exhibit preferably activities related to fungus (Carvalho, 1972). The objective of this research was to verify how the partial removal of the gardening subclass affects two of the activities that occur in the fungus: gardening and care of the brood (eggs, larvae and pupae). The data obtained showed that under conditions that cause a decrease in the number of workers of the gardening subclass, the workers of the neighboring subclasses increase their gardening activity without reducing care of the brood.
Index terms: Animal division of labor. Ants. Age differences. Body size. Animal foraging behavior.
Desde o início deste século o fenômeno do polietismo - tanto em função da idade quanto de características corporais da obreira - tem atraído o interesse de estudiosos do comportamento de insetos sociais (Wheeler, 1910; Buckingham, 1911). A literatura mais recente mostra que esse fenômeno continua sendo alvo de investigação intensa (Vide, p. ex., Wilson, 1971; Carvalho, 1972; Lenoir, 1979; Seeley, 1982, 1983; McDonald & Topoff, 1985, Gerber, Badertscher & Leuthold, 1988).
Carvalho (1972) e Wilson (1980a, 1980b) constataram a existência de divisão de trabalho, em função do tamanho corporal e da idade, entre obreiras de Atta sexdens. Os dados indicaram que a probabilidade de determinada obreira se engajar em uma dada atividade da colônia depende de seu tamanho e idade. Como enfatizou Carvalho (1972), a divisão de trabalho entre as várias subclasses de tamanho/idade é quantitativa e não qualitativa. Isto significa que determinada subclasse se dedica mais a certa atividade, mas também, embora com menor frequência, a outras atividades da colônia.
Wilson (1983) removeu 49% das obreiras forrageadoras de colônias de Atta cephalotes e verificou aumento na atividade forrageadora das 51% remanescentes na colônia. Além disso, as demais subclasses (castas, na nomenclatura adotada por esse autor) não se engajaram em forrageamento. Esse autor interpretou esses resultados como indicadores de que algumas das atividades da colônia de Atta demandam especializações da subclasse de obreiras e, por isso, não podem ser executadas por outras subclasses. Essa conclusão parece incompatível com os dados obtidos por Carvalho (1972) no que se refere à possibilidade de mais de uma subclasse executar a mesma atividade.
O não engajamento de outras subclasses no forrageamento também poderia ser explicado pelo fato de as 51% obreiras forrageadoras terem mantido o forrageamento num nível suficiente. Ou seja, o total de obreiras removido foi insuficiente para produzir ajustamentos tais como a participação de outras subclasses. Tal possibilidade poderia ser testada através da remoção de subclasse envolvida em atividade considerada especializada.
Os gêneros Atta e Acromyrmex são considerados os mais evoluídos dentre todos os gêneros de formigas (Wilson, 1980a, 1980b). Seu alto grau de adaptação pode ser atestado pelo grande número de colônias e capacidade de sobrevivência nas regiões habitadas por essa tribo (Mariconi, 1970). Grande parte de seu potencial adaptativo se deve ao fato de essas formigas cultivarem o seu próprio alimento. De acordo com Wilson (1980a), a tribo Attini foi a única, dentre todas as tribos de formigas, que evoluiu de uma fase meramente coletora-caçadora para o modo de vida agricultor.
Cultivar fungo envolve uma série grande e complexa de atividades, tais como: podar, transplantar, manipular o substrato (vegetais) onde o fungo crescerá, umedecer o fungo. Além dessas operações, as formigas devem também identificar parasitas que ameaçam o fungo (certos tipos de ácaros, por exemplo) e substâncias tóxicas que possam afetá-lo.
Em um estudo sobre as reações de colônias de Acromyrmex a substâncias tóxicas, Machado et al. (1988) verificaram a ocorrência das mesmas reações observadas por outros autores em Atta, tais como: abandono dos olheiros contaminados, remoção de obreiras mortas e vivas contaminadas, remoção de pedaços de esponjas de fungo e mudança da colônia para outro local. Os autores supõem que no decurso da evolução do mutualismo entre as formigas cortadeiras e os fungos por elas cultivados ocorreram pressões seletivas para o desenvolvimento de sistemas e/ou estratégias adequadas à percepção e remoção de agentes capazes de colocarem em perigo a cultura de fungo.
De acordo com Carvalho (1972), as obreiras de no máximo 3 mm de tamanho corporal são as jardineiras por excelência nas colônias de Atta sexdens. O que aconteceria se as obreiras dessa subclasse fossem removidas? A resposta a questões desse tipo, segundo essa autora, traria esclarecimentos importantes acerca da natureza da aparente relação entre tamanho e polietismo. O fato de essa autora ter realizado suas observações em colônias com milhares de formigas impossibilitou a realização de manipulações experimentais que permitissem a remoção de determinados grupos de obreiras.
A presente pesquisa procurou aprofundar a investigação do polietismo, utilizando condições experimentais que permitiam a remoção de obreiras da subclasse jardineira.
Métodos e resultados
Os resultados de alguns de nossos estudos (Rodrigues, 1983; Rodrigues & Ribeiro, 1984) mostraram que é possível utilizar grupos grandes de obreiras como simuladores das atividades da colônia. O procedimento básico desses estudos consistia em retirar da colônia uma porção de esponja de fungo - contendo obreiras e formas imaturas - e acondicioná-la em um recipiente onde as obreiras tivessem livre acesso a vegetais.
O acompanhamento diário desses grupos grandes (com mais de 200 obreiras) permitiu constatar que grande parte das atividades normalmente desenvolvidas na colônia também ocorriam nesses grupos. Em função desses resultados, decidiu-se utilizar esse tipo de preparação na presente pesquisa porque, além das facilidades práticas, essa preparação também facilitaria manipulações experimentais referentes à remoção de obreiras.
Preparação das caixas experimentais. Nos três experimentos realizados, o procedimento básico de preparação consistiu em retirar da colônia, mantida em laboratório, uma porção de esponja de fungo (de aproximadamente 30 cm³), contendo obreiras e formas imaturas. Essa porção de esponja era colocada em um recipiente de vidro de 6 cm de altura por 11 cm de diâmetro, com tampa encaixável. Esse recipiente, com a tampa parcialmente aberta, era colocado em uma bandeja, que tinha as bordas engraxadas para impedir a fuga das obreiras. Na bandeja eram colocados vegetais que as obreiras podiam cortar e transportar para o fungo. Além das obreiras que estavam na porção de esponja, cada caixa recebia também 50 obreiras que haviam sido coletadas na área externa da colônia.
Classificação de tamanho e idade. Para classificar as obreiras em subclasses de tamanho e idade foi utilizada uma adaptação da classificação proposta por Carvalho (1972), que discrimina 10 classes de tamanho e 5 classes de idade (cor).
Tamanho corporal, em nossa classificação, é a medida de comprimento que vai da extremidade distal da mandíbula até à extremidade distal do gastro. A classificação que utilizamos em nossos experimentos foi a seguinte:

A obreira era enquadrada na subclasse 1 quando seu tamanho corporal era menor ou igual a 3 mm; se o tamanho da obreira era maior ou igual a 12 mm, ela era incluída na subclasse 10. Os valores (em mm) das subclasses 2 a 9 são os pontos médios. No caso dessas subclasses, a obreira era enquadrada em determinada subclasse quando seu tamanho corporal se aproximava do ponto médio estabelecido para essa subclasse.
Para classificar a obreira quanto à idade utilizamos a classificação proposta por Carvalho (1972), que estabelece 5 subclasses que vão do amarelo claro (subclasse A) até o marrom escuro (subclasse E), passando por 3 tonalidades avermelhadas correspondentes às subclasses B, C e D.
Delineamento básico e registro dos dados. Em cada experimento foram planejadas duas fases - pré e pós-remoção de obreiras - que duravam no mínimo 5 dias cada uma. Os registros eram realizados duas vêzes ao dia e consistiam em anotar as subclasses de idade e tamanho das obreiras que estavam envolvidas em atividades relativas ao fungo (jardinagem) e às formas imaturas (babás). Ao final do experimento foram registrados os totais de obreiras por subclasse de tamanho/idade de cada caixa experimental. O número de cada tipo de forma imatura (ôvo, larva, pupa) de cada caixa experimental era também computado.
Uma vez que era inviável medir a obreira durante o registro de dados, utilizou-se um cartão onde estavam coladas obreiras representativas de cada subclasse de idade e tamanho. A obreira era então comparada com os modelos e, quando não se conseguia enquadrar a obreira observada em apenas uma das subclasses de tamanho/idade, optou-se por não registrar o evento. Antes do início dos experimentos fizemos um treino de observação e registro que se constituiu no seguinte: apontávamos para um local da colônia e classificávamos a obreira escolhida quanto à idade e tamanho. Em seguida a obreira era capturada e mensurada. O número de êrros e acertos por subclasse foi sendo registrado e o treinamento prosseguiu até que a porcentagem de acertos em relação a qualquer subclasse atingiu no mínimo 90%.
Atividades registradas. Considerava-se cuidar do fungo quaisquer dos seguintes comportamentos das obreiras: implantar vegetal na esponja, implantar fungo na esponja, mastigar vegetal, arrancar (podar) fungo, lamber fungo, transportar fungo, transportar vegetal mastigado. Os cuidados com formas imaturas se referiam a qualquer um dos seguintes comportamentos a elas dirigidos: lamber, alimentar (larva), transportar, esticar partes do corpo da pupa.
Remoção de obreiras. Apenas as obreiras da subclasse de tamanho 1, independente da idade da obreira, foram removidas ao final da Fase 1 dos experimentos. As remoções da subclasse 1 foram parciais em razão da dificuldade de capturar as obreiras que permaneciam no interior da esponja de fungo.
Tratamento dos dados. As frequências das atividades relativas ao fungo e às formas imaturas e o número de obreiras por subclasse de tamanho e idade foram transformados em porcentagens. Para determinar a significância das diferenças entre porcentagens foi consultada a Tábua de Limites Exatos de Confiança para Porcentagens, com nível de significância igual a 0,05 (Geigy, 1965). Optamos por esse método de análise porque foi o utilizado por Carvalho (1972) e, além de se adequar à natureza dos dados, também facilita a comparação de nossos resultados com os obtidos por essa autora.
Os sinais (+,-) que aparecem nas Tabelas significam que o desempenho daquela subclasse na atividade em questão foi considerado significativamente superior (+) ou inferior (-) ao esperado, a partir da consulta à Tábua de Limites. Quando o resultado não vem acompanhado de sinal significa que o desempenho da subclasse naquela atividade ocorreu dentro do esperado. Apesar de os dados terem sido transformados em porcentagens para permitir a consulta da Tábua de Limites, optamos por apresentar nas Tabelas os dados brutos.
Experimentos R1
Método
Foram preparadas 10 caixas experimentais que foram acompanhadas no decorrer de 10 dias. Findos os primeiros 5 dias (Fase 1), procedeu-se à remoção parcial da subclasse 1 e acompanhamento das caixas por mais 5 dias (Fase 2).
Resultados
Ao final da Fase 1 foram removidas 1296 obreiras da subclasse de tamanho 1, o que representa 61,71% do total de obreiras dessa subclasse. A despeito de a subclasse 1 ter sido reduzida em mais de 50%, os resultados mostram que essa subclasse ainda manteve a supremacia do trabalho no fungo na Fase 2 (Vide Tabela 1).

Embora tenha havido diminuição da jardinagem realizada pela subclasse 1 após a remoção parcial, o total de jardinagem exibido na Fase 2 ainda foi significativamente maior que o esperado. Nos cuidados com formas imaturas, por outro lado, o trabalho dessa subclasse, que já era significativamente abaixo do esperado, sofreu uma grande redução na Fase 2.
A Tabela 1 também mostra que as subclasses 2 e 3 apresentaram modificações no seu padrão de desempenho. Em relação à jardinagem, na Fase 1 a subclasse 2 mostrou desempenho de acordo com o esperado. Na Fase 2 ocorreu uma inversão no desempenho dessas duas subclasses, tanto em jardinagem quanto nos cuidados com formas imaturas. Apesar de a Tabela 1 indicar essa inversão de papéis das subclasses 2 e 3, entre as duas fases do experimento, note-se que a magnitude dessa inversão é diferente para as duas atividades. Em jardinagem o desempenho das obreiras 2 e 3 oscilou de significativamente abaixo, até o esperado. No trabalho com as formas imaturas, por outro lado, o intervalo de oscilação vai de significativamente acima, até de acordo com o esperado. Isto pode estar indicando uma substituição em cadeia, onde a subclasse 2 compensa parcialmente o trabalho em jardinagem que deixou de ser realizado em função da remoção parcial da subclasse 1, enquanto a subclasse 3 compensa a ausência parcial da subclasse 2 nos cuidados de formas imaturas.
Quanto ao desempenho em função da idade (Tabela 2), verificou-se que a subclasse C foi a única a exibir trabalho significativo na Fase 2, tanto em jardinagem como nos cuidados com formas imaturas. Pode-se inferir, a partir da análise das Tabelas 1 e 2, que as subclasses 1C e 2C foram as principais responsáveis pelo trabalho no fungo na Fase 2.

Experimento R3
Os resultados do experimento anterior mostraram que, a despeito da remoção de mais de 50% da subclasse de tamanho 1, o total de jardinagem exibida por obreiras remanescentes dessa subclasse permaneceu significativamente alto. Como a subclasse 1 era a mais numerosa, ainda restou um contingente de 804 obreiras após a remoção, número esse suficientemente alto para permitir novos ajustamentos às atividades dentro dessa subclasse. Se o número de obreiras de tamanho 1 fosse reduzido ainda mais, isto acarretaria ajustamentos que envolveriam mudanças mais radicais nos padrões das outras subclasses? Este experimento foi planejado com o objetivo de testar essa hipótese.
O método utilizado para remover as obreiras das caixas experimentais não permitia retirar as obreiras que se encontravam no interior da esponja. Quando se iniciava a operação de remoção, as obreiras tendiam a se refugiar no interior da esponja. Quando as perturbações provocadas pelo procedimento de remoção cessavam, as obreiras saíam dos esconderijos e retomavam as atividades. Para possibilitar a remoção do maior número possível de obreiras da subclasse 1, o procedimento de remoção foi repetido três dias consecutivos.
Método
Foram preparadas 5 caixas experimentais semelhantes às do experimento anterior. O procedimento consistiu em duas fases, sendo a Fase 1 idêntica à do experimento anterior. Ao término da Fase 1 foi feita a primeira remoção de obreiras da subclasse 1. O procedimento de remoção dessa subclasse foi repetido nos dois dias subseqüentes, registrando-se o número retirado de cada caixa. Decorridas 3 a 5 horas desde cada uma das remoções, realizava-se a primeira observação do desempenho das obreiras nas atividades de interesse; a segunda observação do dia era realizada no final da tarde. As caixas experimentais foram mantidas por 5 dias após a última remoção de obreiras (Fase 2), registrando-se os eventos do mesmo modo que na Fase 1.
Resultados
A repetição do procedimento de retirada permitiu remover cerca de 80% (1499) das obreiras presentes na Fase 1. A Tabela 3 apresenta os dados da Fase 1 e os dos 5 dias posteriores à última remoção de obreiras (Fase 2). Apesar de a subclasse 1 continuar apresentando os maiores totais de jardinagem, quando comparada com as demais subclasses, seu desempenho na Fase 1 foi considerado significativamente menor do que o esperado.

Assim, enquanto a subclasse 1 apresenta desempenho em jardinagem significativamente baixo, as subclasses 2, 3 e 4 apresentam desempenho significativamente superior ao esperado. Talvez se possa inferir que quando, por alguma razão, a subclasse 1 está pouco envolvida com jardinagem, as subclasses adjacentes se dedicam mais a essa atividade.
O baixo desempenho em jardinagem da subclasse 1 na Fase 1, apresentado na Tabela 3, foi devido ao baixo desempenho exibido pelas obreiras de duas caixas experimentais nos três primeiros dias da Fase 1. Não conseguimos saber o que poderia ter produzido tal efeito.
A tabela 4 foi construída com o objetivo de avaliar a rapidez do ajustamento das obreiras às situações de remoção. Nessa Tabela são apresentados os totais do último dia da Fase 1, de cada um dos três dias de remoção e do último dia da Fase 2. Os totais de jardinagem e cuidados com formas imaturas foram obtidos somando-se os dados nas duas observações diárias de todas as caixas.

Como se pode observar na Tabela 4, a jardinagem da subclasse 1 se apresentou dentro do esperado no último dia da Fase 1, e se manteve no mesmo nível após cada retirada (nos três dias foram removidas 344, 314 e 263 obreiras, respectivamente). No último dia da Fase 2, decorridos 4 dias desde a última remoção de obreiras, a subclasse 1 exibiu desempenho como jardineira significativamente maior que o esperado. Quanto ao desempenho como babá, a subclasse 1 se manteve sempre abaixo do esperado, enquanto a subclasse 2 apresentou desempenho significativamente alto até a segunda remoção.
Os resultados da Tabela 4 mostraram que, de um modo geral, a subclasse 1 e as demais obreiras mantiveram o mesmo padrão de trabalho no fungo e com as formas imaturas, a despeito da retirada de obreiras. Isto sugere que a redistribuição das obreiras remanescentes pelas atividades é muito rápida, uma vez que as primeiras observações foram feitas 3 a 5 horas após a remoção das obreiras.
Quanto à idade, verifica-se na Tabela 5 que as obreiras da subclasse C foram as únicas a apresentarem desempenho significativamente superior ao esperado como jardineiras e babás, em ambas as fases. As subclasses B e D apresentaram desempenho abaixo do esperado. A subclasse A só esteve presente na Fase 2 - porque ocorreram eclosões na Fase 2 - e em número muito baixo.

Experimento R4
Os resultados dos experimentos R1 e R3 apontaram na direção da existência de ajustamentos que envolviam substituição dentro da subclasse de tamanho 1. Isto significa que na ausência do grupo trabalhador outras obreiras de mesmo tamanho, mas menos dedicadas à jardinagem, passam a trabalhar no fungo de modo intensivo. O que aconteceria se as obreiras removidas ao final da Fase 1 retornassem às suas caixas? O desempenho da subclasse 1 em jardinagem aumentaria ou tenderia a se estabilizar? Este experimento foi planejado para tentar responder essas questões.
Método
Foram preparadas 5 caixas experimentais similares às dos experimentos anteriores. Foram planejadas três fases, sendo as Fases 1 e 2 semelhantes às dos experimentos anteriores. Ao final da Fase 1 procedia-se a remoção das obreiras da subclasse 1 que, ao invés de serem eliminadas como ocorria nos experimentos anteriores, eram colocadas em caixas experimentais contendo água e fragmentos de esponjas de fungo que serviam como alimento.
As obreiras removidas de cada caixa experimental eram contadas e permaneciam nessa nova caixa durante toda a Fase 2, sendo devolvidas à sua caixa de origem antes do início da Fase 3.
As técnicas de observação e registro, bem como os demais passos do procedimento, foram idênticos aos dos experimentos anteriores.
Resultados
Como a remoção de obreiras foi feita uma única vez, o total de obreiras removidas foi igual a 737 (cerca de 47%). Repetiu-se o curioso resultado obtido no experimento anterior, onde as obreiras de tamanho 1 apresentaram, na Fase 1, desempenho em jardinagem significativamente inferior ao esperado (Vide Tabela 6). Na Fase 2, quando 47% da subclasse 1 estava ausente, o trabalho no fungo aumentou. A volta das obreiras retiradas (Fase 3) fez aumentar o total de trabalho no fungo.

O ritmo de trabalho nas três fases pode ser melhor avaliado observando-se os resultados da Tabela 7, que apresenta os totais dos dois últimos e dois primeiros dias de cada fase. O desempenho em jardinagem da subclasse 1 se manteve dentro do esperado no final da Fase 1 e no início da Fase 2, o que pode indicar que as obreiras removidas foram imediatamente substituídas por suas congêneres de tamanho. Ao final da Fase 2 o desempenho das "substitutas" chegou a ser significativamente maior que o esperado, o que é indicativo de aumento significativo na performance dessas obreiras. A manutenção do desempenho dentro do esperado, no início da Fase 3, leva à suposição de que parte das obreiras que retornaram às caixas se engajaram em jardinagem, e esse engajamento deve ter aumentado porque o trabalho no fungo tornou-se significativamente maior que o esperado, nos últimos dois dias do experimento.
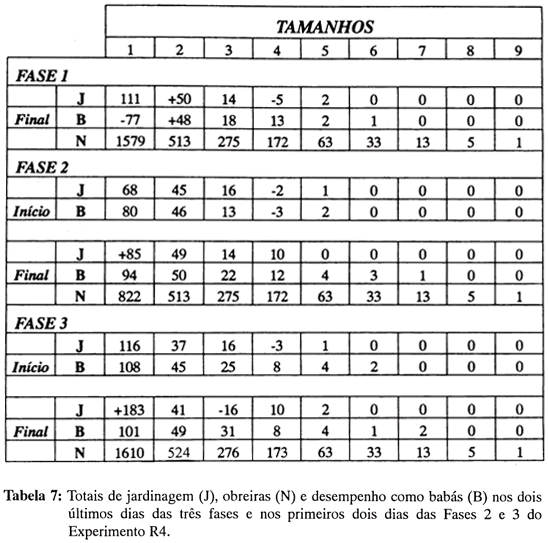
Ainda no que se refere à subclasse 1, verificou-se que os cuidados com formas imaturas aumentaram da Fase 1 para a Fase 2, ficando significativamente acima do esperado. Dentre as demais subclasses de tamanho, a única que merece destaque é a de número 2, que exibiu na Fase 1 desempenho significativo em relação ao fungo e às formas imaturas. Essas obreiras mantiveram praticamente o mesmo desempenho na Fase 2 - embora já não sendo considerado significativo - o que indica que o comportamento dessa subclasse não foi afetado pela retirada ou pela volta das obreiras de tamanho 1.
O desempenho da subclasse de idade C em jardinagem (conforme Tabela 8) manteve-se acima do esperado nas três fases do experimento, enquanto as subclasses A, B e D apresentaram, de modo geral, trabalho no fungo significativamente abaixo do esperado. Nos cuidados de formas imaturas, por outro lado, o desempenho da subclasse C só foi considerado estatisticamente significativo na Fase 1. Já as subclasses de obreiras mais jovens (A e B) apresentaram desempenhos como babás que chegaram a ser considerados significativamente acima do esperado.

Discussão
A comparação entre os nossos resultados das Fases 1 e os de Carvalho (1972), obtidos através de observações de colônias intactas, mostra semelhanças entre eles. Dentre as similaridades destacamos: (1) predominância da subclasse 1 no trabalho de jardinagem e decréscimo de desempenho nessa atividade com o aumento do tamanho corporal; (2) predominância nos cuidados de formas imaturas das subclasses imediatamente superiores à subclasse 1. Tais semelhanças ajudam a corroborar a validade do procedimento e das condições experimentais utilizados em nossos experimentos.
A discrepância mais proeminente entre os resultados desta pesquisa e os obtidos por Carvalho (1972) é referente à subclasse de idade mais freqüente na esponja de fungos. Essa autora verificou que a subclasse B era a de presença mais expressiva, enquanto nossos resultados mostraram que C era a mais numerosa. O local da esponja de fungo da colônia de onde retirávamos as porções de esponja pode explicar essa discrepância. Sempre retirávamos as porções de esponjas para os experimentos de local na colônia onde o fungo era novo. É possível que os locais onde o fungo está sendo plantado congreguem obreiras mais velhas, já que nesse local ocorrem também certas atividades relacionadas à preparação do vegetal - mascar, cortar - que são executadas por obreiras de idade mais intermediária.
Nossos resultados mostraram que, além de mais numerosa, a subclasse C também apresentou grande participação em jardinagem. De acordo com Carvalho (1972) quando a obreira de idade intermediária (C) pertence à subclasse 1 ocorre algum tipo de interação entre idade e tamanho que faz dessa obreira uma excelente jardineira. Esse efeito explicaria o porque do alto desempenho da subclasse C em jardinagem; a grande maioria dessas obreiras pertenciam também à subclasse 1.
Os resultados das Fases 2 mostraram que as obreiras das subclasses de tamanho próximas (2, 3 e 4) exibiram mais atividades de jardinagem após a remoção parcial da subclasse 1. No entanto, essa substituição, relativa à jardinagem, exibida pelas obreiras das subclasses próximas, foi fraca. Os efeitos mais notáveis da remoção parcial foram: (1) aumento significativo na atividade jardineira e diminuição do cuidar de formas imaturas pelas obreiras 1 remanescentes; (2) aumento significativo nas atividades relativas às formas imaturas exibido pelas obreiras das subclasses de tamanho 2 e 3. Esses resultados levam à dedução de que o efeito mais proeminente de substituição pelas subclasses próximas ocorreu em uma atividade secundária da subclasse 1 - o cuidar de formas imaturas.
Remoções parciais de obreiras forrageadoras efetuadas por Kolmes e Winston (1988) e Wilson (1983) em colônia de Apis mellifera e Atta cephalotes, respectivamente, também não produziram substituições significativas por parte das subclasses próximas. Kolmes e Winston (1988) concluíram que a ausência de alterações significativas em polietismo foi devida ao fato de as manipulações serem brandas; somente manipulações drásticas - tais como, remoção de todas as obreiras forrageadoras - produziriam alterações significativas em polietismo. Wilson (1983), por outro lado, interpretou a ausência de substituição como prova de que no gênero Atta a atividade forrageadora pertence ao conjunto de atividades que o autor denominou fixas, ou seja, exigem certas especializações que somente determinada subclasse possui (Wilson, 1980a).
Diversos estudos citados por Kolmes e Winston (1988) mostram que, a despeito de o polietismo etário ser uma regra geral em colônias de abelhas, existe grande variabilidade individual. Os autores interpretam essa variabilidade como uma manifestação das colônias de abelhas a diferentes pressões ambientais e ergonômicas. Quando as pressões são brandas, não ocorrem mudanças nas idades em que normalmente as obreiras executam determinadas tarefas; ou seja, não há alteração no polietismo etário. Sob condições de manipulações drásticas, o polietismo etário deixa de existir - pelo menos temporariamente. Os autores supõem então que as colônias de abelhas são sistemas com grande elasticidade, nos quais existe grande margem para pequenas mudanças e adaptações antes que sejam necessárias mudanças drásticas no padrão geral da colônia.
Acreditamos que nossos experimentos, bem como a pesquisa de Wilson (1983), também podem ser considerados exemplos de manipulações brandas e que aos resultados obtidos se aplicariam as teorizações propostas por Kolmes e Winston (1988). As substituições pelas obreiras remanescentes da subclasse removida, a rapidez da adaptação pós-remoção e a substituição pelas subclasses adjacentes em atividade secundária (cuidar de formas imaturas) da subclasse 1, podem estar revelando a elasticidade do sistema (colônia) e suas possibilidades de adaptação.
A variabilidade individual, considerada por esses autores como o fator que tornaria possíveis as adaptações da colônia, tem estado presente em todas as nossas pesquisas com Atta sexdens (Vide, p. ex., Rodrigues, 1983; Rodrigues & Ribeiro, 1984; Rodrigues & Ribeiro, 1991). Dentre os autores que relataram a existência de diferenças individuais em formigas, podemos citar as pesquisas de Chen (1937) com Camponotus japonicus, Moglich (1978) com Leptothorax e Morley (1954) com Formica rufa na escavação, migração do ninho e aprendizagem, respectivamente.
Schmidek et al. (1991), analisando a ocorrência de diferenças individuais no desempenho de roedores, concluiram que essas diferenças têm raízes filogenéticas e, além disso, provavelmente se relacionam com a organização social. Pode-se deduzir a partir das conclusões desses autores que a variação individual é própria das espécies sociais. Cabe ressaltar que a variabilidade individual não é incompatível com a especialização; ao contrário, a possibilidade de variação acrescenta maleabilidade a um sistema onde a divisão de trabalho é bem estabelecida. Em síntese, pode-se afirmar que a variabilidade individual permitiria à colônia (ou grupo) a adaptação a mudanças (ou pressões), mantendo os seus padrões de divisão de trabalho.
Se aceitarmos que as manipulações realizadas nesta pesquisa e na realizada por Wilson (1983) são manipulações brandas, resta saber o que aconteceria se as pressões sobre a colônia fossem drásticas. Ou seja, quanto de pressão uma colônia de Atta pode suportar mediante os ajustamentos decorrentes da variabilidade individual?
REFERÊNCIAS BIBLIOGRÁFICAS
BUCKINGHAM, E.N. Division of labor among ants. Proceedings of the American Academy of Arts and Sciences, v.46, n.18, p.426-507, 1991. [ Links ]
CARVALHO, A.M.A. Alguns dados sobre a divisão de trabalho entre obreiras de Atta sexdens rubropilosa Forel, 1908 (Hymenoptera-Formicidae), em colônias iniciais, mantidas em laboratório. São Paulo, 1972. 175p. Tese (Doutorado) - Instituto de Psicologia, Universidade de São Paulo. [ Links ]
CHEN, S.C. The leaders and followers among the ant in nest-building. Physiological Zoology, v.10, p.437-55, 1937. [ Links ]
GEIGY, J.R, ed. Tablas científicas. Basilea, J.R. Geigy, 1965. [ Links ]
GERBER, C.; BADERTSCHER, S.; LEUTHOLD, R.H. Polyethism in Macrotermes bellicosus (Isoptera). Insectes Sociaux, v.35, n.3, p.226-40, 1988. [ Links ]
KOLMES, S.A.; WINSTON, M.L. Division of labor among worker bees in demographically manipuled colonies. Insectes Sociaux, v.35, n.3, p.262-70, 1988. [ Links ]
LENOIR, A. Feeding behaviour in young societies of the ant Tapinoma erraticum L.: trophallaxis and polyethism. Insectes Sociaux, v.26, n.1, p.19-36, 1979. [ Links ]
McDONALD, P.; TOPOFF, H. Social regulation of behavioral development in the ant Novomessor albisetsus (Mayr). Journal of Comparative Psychology, v.99, n.1, p.3-14, 1985. [ Links ]
MACHADO, V.; DIEHL-FLIEG, E.; SILVA, M.E.; LUCCHESE, M.E.P. Reações observadas em colônias de algumas espécies de Acromurmex (Hymenoptera-Formicidae) quando inoculadas com fungos entomopatogênicos. Ciência e Cultura, v.40, n.11, p.1106-8, 1988. [ Links ]
MARICONI, F.A.M. As saúvas. São Paulo, Ceres, 1970. [ Links ]
MOGLICH, M. Social organization of the nest emigration in Leptothorax (Hymenoptera-Formicidae). Insectes Sociaux, v.25, n.3, p.205-25, 1978. [ Links ]
MORLEY, D.W. The evolution of an insect society. London, George Allen & Unwin, 1954. [ Links ]
RODRIGUES, M.M.P. Estudo dos fatores sociais envolvidos na escavação do solo por obreiras de Atta sexdens rubropilosa Forel, 1908 (Hymenoptera-Formicidae). São Paulo, 1983. 124p. Dissertação (Mestrado) - Instituto de Psicologia, Universidade de São Paulo. [ Links ]
RODRIGUES, M.M.P.; RIBEIRO, F.L. Construção do ninho por obreiras de Atta sexdens. Ciência e Cultura, v.36, n.7, p.972, 1984. Suplemento. /Apresentado à 36. Reunião Anual da SBPC, São Paulo, 1984 - Resumo/ [ Links ]
RODRIGUES, M.M.P.; RIBEIRO, F.L. Facilitação social na escavação do solo por pequenos grupos de obreiras Atta sexdens rubropilosa. Biotemas, v.4, n.1, p.63-74, 1991. [ Links ]
SCHMIDEK, W.R.; NISHIDA, S.M.; PINTO, C.M.H.; SCHMIDEK, M. Diferenças individuais no comportamento de roedores. Biotemas, v.4, n.1, p.53-61, 1991. [ Links ]
SEELEY, T.D. Adaptive significance of the age polyethism schedule in honeybee colonies. Behavioral Ecology and Sociobiology, v.11, p.287-93, 1982. [ Links ]
SEELEY, T.D. Division of labor between scouts and recruits in honeybee foraging. Behavioral Ecology and Sociobiology, v.12, p.253-9, 1983. [ Links ]
WHEELER, W.M. The ants: their structure, development and behavior. New York, Columbia University Press, 1910. [ Links ]
WILSON, E.O. Caste and division of labor in leaf-cutter ants (Hymenoptera-Formicidae: Atta). I. The overall pattern in Atta sexdens. Behavioral Ecology and Sociobiology, v.7, p.143-56, 1980a. [ Links ]
WILSON, E.O. Caste and division of labour in leaf-cutter ants (Hymenoptera-Formicidae: Atta). II. The ergonomic optimization of leaf cutting. Behavioral Ecology and Sociobiology, v.7, 157-65, 1980b. [ Links ]
WILSON, E.O. Caste and division of labour in leaf-cutter ants (Hymenoptera-Formicidae: Atta). III. Ergonomic resiliency in foraging by A. cephalotes. Behavioral Ecology and Sociobioloy, v.14, p.47-54, 1983. [ Links ]
WILSON, E.O. The insect societies. Cambridge, Harvard University Press, 1971. [ Links ]
1 Parte da tese de Doutorado em Psicologia - área de concentração: Psicologia Experimental - IPUSP, 1989. Orientador: Vera Silvia Raad Bussab.
