










Serviços Personalizados
artigo
 Português (pdf)
Português (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
Indicadores

Compartilhar

 Permalink
PermalinkPsicologia USP
versão On-line ISSN 1678-5177
Psicol. USP v.6 n.1 São Paulo 1995
ARTIGOS ORIGINAIS
A construção da teia geométrica enquanto instinto: primeira parte de um argumento1
Orb-web building as an instinct: the first part of an argument
César Ades
Instituto de Psicologia - USP
RESUMO
Entende-se geralmente a construção da teia orbicular como um comportamento estereotipado e resistente à aprendizagem. A fim de pôr à prova esta idéia e de detectar possíveis efeitos de experiência, aranhas orbitelas adultas, Argiope argentata (Fabricius), foram mantidas por um período prolongado em caixas-suportes de diferentes tamanhos, sendo assim levadas a construir teias de proporções e tamanhos diversos. Eram depois testadas numa caixa-suporte de tamanho intermediário. Supunha-se que a experiência prévia em tecer em contextos maiores ou menores se traduziria por diferenças nas proporções internas e/ou no tamanho das teias da fase de teste. Os dados não corroboraram esta expectativa. Podem ser tomados os resultados como mais um argumento a favor de uma visão instintivista da construção da teia: prefiro tomá-los como uma indicação de que alguns aspectos da história passada não afetam as características básicas e flexíveis do programa comportamental de construção. A questão do papel que a aprendizagem pode desempenhar na habilidade tecedora da aranha permanece aberta.
Descritores: Comportamento instintivo animal. Aracnídeos. Construção da teia. Aprendizagem animal. Etologia animal.
ABSTRACT
Orb-web building is usually interpreted as a stereotyped, resistant-to-learning performance. In order to detect eventual effects of experience on web building, groups of adult orb-web spiders Argiope argentata (Fabricius) were kept for an extended period in frames of different dimensions, spinning different sized webs. They were then tested in a frame of an intermediate size. It was hypothesized that previous experience of building in larger or smaller contexts would have effects on the size or internal proportions of the test webs. Data did not corroborate such an expectation: results may be taken as reinforcing an instinctivist point of view: I prefer to interpret them as showing that some aspects of past experience do not affect the basic features of the web-building program. The question of the role learning may play in the spider's spinning skills remains open.
Index Terms: Animal instinctive behavior. Arachnida. Web construction. Animal learning. Animal ethology.
A aranha é que é bacana
com sua geometria
Euclidiana
Millôr Fernandes - Hai-Kais
A geometria da teia de uma aranha orbitela nasce aos poucos, de pormenor em pormenor, produto da execução de atos repetitivos cuja natureza muda de uma etapa a outra, dentro da programação geral de construção. Se a teia é euclidiana, é mercê da aplicação de regras comportamentais que se integram de forma hierárquica, controlando desde o modo como um fio é grudado a outro até as trajetórias de locomoção através das quais são postas as espirais (Ades, 1986; Eberhard, 1972; Peters, 1936; Ramousse, 1982; Tilquin, 1942; Vollrath, 1992; Witt, Reed & Peakall, 1968).
De modo bastante original e sugestivo, Vollrath (1992) interpreta a teia da aranha como "(...) uma trajetória de forrageamento que se congelou (...) uma trajetória de busca que foi terminada muito antes de qualquer presa ser capturada, uma reação filogenética, não uma reação imediata, à presa" (p.147).
É uma busca ritualizada das muitas presas que a aranha aceita como alimento, da presa em geral. Começa com uma locomoção exploratória por folhas e hastes, suportes potenciais, durante a qual a aranha poderia estar colhendo informações espaciais e outras informações importantes para a escolha do local de construção - alturas, distâncias livres, pontos de amarração, direção do vento etc. De que modos a informação é armazenada e utilizada, como são feitas as estimativas espaciais, não sabemos ainda.
Numa etapa seguinte, são postos, de forma integrada, os fios de amarra, os fios do quadro e os raios. É o arcabouço da teia, todo feito de seda seca, um andaime sobre o qual partes susbseqüentes poderão ser fixadas, uma delimitação do espaço que poderá ser aproveitado pela teia. Em seguida (dizer em seguida é fazer uso de licença para a simplificação da exposição: os comportamentos das duas etapas se entremeiam, em sua fronteira temporal), a aranha constrói uma espiral seca, girando de dentro para fora, espiral esta que, compacta no início, dá origem a voltas bastante espaçadas, a partir de uma certa distância do centro, constituindo então a espiral provisória. Finalmente, trabalhando de fora para dentro, destruindo pari passu a espiral provisória, a aranha coloca a espiral viscosa ou espiral captora, conserta o centro da teia, substituindo seus fios por uma rede irregular de fios e, antes de entrar na postura de repouso (em se tratando de Argiope argentata), coloca de uma a quatro estruturas de seda bem clara, em zigue-zague, mais ou menos na direção das patas, os estabilimenta ou ornatos da teia, que têm por função atrair insetos e de que certos insetos tentam fugir (Craig & Bernard, 1990; Craig, 1994).
Uma vez postas as amarras e uma vez pronto o quadro da teia, o comportamento da aranha passa a depender, basicamente, a cada passo, de uma leitura da estrutura de fios de seda já existente. A lógica da construção é, portanto, uma lógica do acréscimo, a partir de um algoritmo, de elementos a outros elementos cuja disposição e tamanho são levados em conta. A teia é ambiente para a feitura de si-própria, sua flexibilidade de construção depende, além do ajustamento aos suportes, efetuado na etapa de constituição do arcabouço, do ajustamento de cada fio aos fios contextuais.
Seguindo uma argiope, num dos momentos de sua construção da espiral viscosa, vê-se quão delicada é a discriminação do ambiente e quão precisos são os movimentos orientados. A aranha alterna locomoção e colagem do fio viscoso. Durante a locomoção, as patas do primeiro par constantemente monitoram os fios contextuais, tocam de leve na última espira viscosa posta, ou na espiral provisória, que serve de guia (Zschokke, 1993). As patas II e III alternam-se como em qualquer andança, mas as patas II, dispostas muito mais para o lado do que normalmente, acabam tendo (procuram ?) pontos especiais de agarramento, a pata II central num raio, a pata II periférica no raio anterior. A pata IV periférica vem colocar-se perto da pata III ipsilateral (num raio em que haverá colagem) enquanto a pata IV, iniciando a fase de colagem, apanha o fio viscoso em sua saída das fiandeiras e, esticando-se, o estica também e afrouxa. Quanto esta pata atinge o seu máximo de extensão, o abdomen se curva ventralmente em direção ao raio de colagem, ao mesmo tempo em que as patas III e IV periféricas nele agarradas se dobram bruscamente. Graças a esta dupla aproximação, as fiandeiras tocam no raio, nele se esfregam e há colagem. A pata IV solta o fio viscoso e tudo recomeça (Ades, 1972; ver também a descrição da colagem do fio viscoso em Leucauge mariana, Eberhard, 1982).
A concatenação de atos que vemos, a cada unidade do processo iterativo de colocação da espiral viscosa, provavelmente tenha se originado, num passado filogenético difícil de localizar, de adaptações do movimento locomotor simples. Deve estar codificada no sistema nervoso, não de uma forma balística, em encadeamento irresistível de componentes, de acordo com o modelo lorenziano do padrão fixo de respostas, mas de uma forma cibernética, de "busca" de conseqüências e de correção de trajetória (Ades, 1986). Além disso, é possível que nela estejam embutidos processos de memória. Esta é a crença de Eberhard (1988b, comunicação pessoal), a partir de observações do modo como Leucauge mariana reage, durante a construção da espiral provisória, ao atingir pela primeira vez uma espira provisória prévia ou ao passar por uma região onde a tesoura do experimentador aboliu segmentos provisórios prévios. L. mariana usaria representações das distâncias percorridas, no passado imediato, como critério para a determinação dos pontos de colagem.
A discussão acerca da natureza instintiva vs. adquirida da construção, para a qual o presente artigo pretende contribuir, é tão antiga quanto o interesse por teias geométricas. Na verdade, ela quase sempre foi feita em termos favoráveis à hipótese instintivista. A teia geométrica é um exemplo-de-manual clássico (eu também o uso sempre em aula, quando é preciso sacudir a poeira de um ambientalismo extremo). A ela se aplicam os critérios tradicionais : o caráter típico-da-espécie, o surgimento precoce e a resistência a condições drásticas de privação, durante a ontogênese.
O caráter típico-da-espécie. Teias de orbitelas de espécies ou gêneros diferentes podem muitas vezes ser distinguidas pela forma geral, pelo tipo de estabilimentum, pelo refúgio ou ausência dele, pela existência de um setor vazio, pela cor da seda, pelo modo de amarração à vegetação etc., o que tem tornado freqüente a afirmação de que a teia é típica-da-espécie ou típica-do-gênero (Foelix, 1982; Witt, 1968). Características estruturais das teias foram, inclusive, usadas como critérios taxonômicos (Kaston, 1964).
Nem sempre é fácil, contudo, distinguir entre espécies próximas a partir de suas teias; há uma grande variabilidade nas medidas obtidas dentro de cada espécie ou gênero, os parâmetros medidos em teias nem sempre se coadunam com critérios outros de classificação, a interpretação em termos de convergência parece muitas vezes a mais conveniente. Apelos para mais cautela no uso de feições da teia como caracteres taxonômicos têm sido feitos (Levi, 1978; Eberhard, 1990). Uma abordagem mais recente tem, assim mesmo, ressaltado o valor dos padrões motores empregados na colocação dos fios (em contraposição a aspectos de teias prontas) como elementos taxonômicos válidos, por serem "estreitamente controlados pelo programa genético da aranha" (Shear, 1994, p.257).
A primeira teia. Quando passam do estágio larval para o de ninfa, as aranhas orbitelas imaturas ainda permanecem alguns dias na ooteca antes de ganhar o mundo externo onde se aglomeram numa espécie de "teia comunitária". São centenas de filhotes formando uma pelota aproximadamente esférica. Afastam-se coletivamente, quando perturbados, como se inchasse um super-organismo, e se reagrupam em seguida. Nada constróem que tenha a geometria de uma teia. A primeira teia, que surgirá depois de se espalharem, aproveitando o vento e depois de passarem por mais uma muda, é minúscula, porém perfeita. As fotos destas primeiras teias, de Araneus diadematus (Witt, 1971a), Argiope aurantia (Le Guelte, 1968) e outras nos mostram réplicas de teias adultas, dotadas dos elementos essenciais. As teias precoces seriam, segundo alguns relatos, mais regulares do que as de espécimes maduros (Witt, Rawlings & Reed, 1972).
Tenho visto primeiras teias imperfeitas. Ninfas de A. argentata, que eu retirava de sua "teia comunitária" mais ou menos na época do começo da dispersão, e que eu colocava em presença de suportes apropriados, teciam muito rapidamente uma teia geométrica, mas, muitas vezes, com irregularidades, raios que convergiam na periferia, espiras grudadas umas às outras etc. Teias posteriores eram, em geral, melhores. Sem dados de controle que verificassem uma explicação em termos de maturação, eu não poderia contudo falar em aprendizagem. Confesso-me tentado a replicar as observações, para tirar a limpo a questão.
O papel da maturação é evidenciado em observações como a de Peters (1969) em que espécimes imaturos de Z. x-notata eram retirados da ooteca, antes do tempo. Verificou-se que já soltavam um fio guia e que, em alguns casos, teciam um refúgio rudimentar. Alguns dias depois, faziam uma teia com irregularidades e, mais tarde ainda, uma teia perfeita, do jeito que todas as zigielas jovens tecem. Ninfas de Argiope trifasciata também tecem se liberadas prematuramente da ooteca, uma teia com disposição irregular dos raios e com número muito reduzido de espiras viscosas (Le Guelte, 1968). Araneus suspicax, que normalmente faz sua primeira teia depois da segunda muda ninfal, pode ser induzida a tecer antes da muda, se isolada (o agrupamento como que inibe o surgimento da construção). Trata-se, então de uma teia muito pobre, às vezes com apenas duas ou três espiras viscosas, que a aranha, assim mesmo, consegue usar para capturar insetos (Ramousse & Wurdak, 1984). Em todos estas pesquisas, o tempo, e o amadurecimento que ele propicia, parecem suficientes para o aperfeiçoamento da construção e para o surgimento da teia perfeita.
As aranhas orbitelas possuem, assim, nos primórdios de seu desenvolvimento, uma informação sensório-motora necessária para a elaboração de uma teia funcional. Poder-se-ia dizer que está satisfeita a primeira condição para se falar em comportamento instintivo. São poucas, contudo, as pesquisas que tentem manipular as condições ambientais, durante esta fase precoce, verificando se e como mudanças podem ser incorporadas ao algoritmo de construção.
Experimentos de privação. São muito citados (porém, poucas vezes replicados) os experimentos de Kaspar Hauser de Petrusewiczowa (1938) e de Mayer (1953). Ambas as pesquisadoras usaram Zygiella x-notata. Esta aranha, quando jovem, tece teias orbiculares completas e, mais tarde, teias em que um setor é deixado livre de espiras viscosas, sendo atravessado por um fio único que vai do centro a um refúgio externo (uma folha, por exemplo), onde reside então a aranha. Petrusewiczowa e Mayer criaram por três semanas espécimes de Z. x-notata em tubos estreitos de vidro, impedidas de tecer. Soltas, as aranhas produziam, de imediato, a teia sem um dos setores, típica do estágio mais maduro da espécie. Conclui cada uma das autoras que uma experiência de construção não é pré-requisito necessário para este "pulo" ontogenético (Witt, 1968).
No experimento de Reed, Witt, Scarboro e Peakall (1970), em que se buscou manipular, dentro do paradigma de privação, o treino prévio de construir e de caçar, grupos de dezoito aranhas Araneus diadematus, pertencentes à mesma ooteca, foram submetidos cada qual a uma das seguintes condições experimentais: (1) as aranhas eram criadas em tubos de vidro e alimentadas diretamente; (2) as aranhas eram criadas em tubos de vidro, com a possibilidade de caçar em teias alheias; (3) as aranhas teciam teias mas eram alimentadas diretamente (nunca caçavam) e (4) as aranhas eram livres de tecer e de caçar em suas próprias teias. Testadas, 25 semanas depois, num ambiente padronizado, todas conseguiram construir teias funcionais. A experiência de caçar em teia própria ou em teia alheia, não teve impacto: as aranhas "caçadoras" e as alimentadas na boca construíam teias similares. A criação em tubos, contudo, levou à redução do tamanho da teia, um efeito que desapareceu, passada uma semana.
Estes resultados revelam, de um lado, a estereotipia do desempenho, sua independência em relação a eventos talvez relevantes (o uso da teia como instrumento de caça); de outro, indicam, ao contrário, que o desempenho sofre mudança, em função da história do organismo (da falta de oportunidade de tecer). Desvendam, a um tempo, aspectos instintivos e plasticidade. Uma ressalva: a mudança de tamanho da teia, em aranhas privadas, não se deve necessariamente a um mecanismo comportamental. Submetidas a uma menor demanda ambiental, as aranhas poderiam ter desacelerado sua produção de seda: a redução da teia seria indício de estoques menores nas glândulas sericígenas.
O fator "gasto de seda" foi parcialmente controlado num trabalho posterior da equipe de Witt (Witt, Reed & Scarboro, 1969), do qual somente pude me inteirar a partir de um resumo de comunicação científica. Um grupo de aranhas criadas em tubos, por 35 ou 94 dias, teve seu desempenho comparado, depois de soltas, ao de aranhas, de mesma idade e mesmo peso, criadas sem restrição. As primeiras teias das aranhas experimentais diferiam significativamente, numa série de parâmetros, em ambos os prazos de soltura, das teias-controle. O esvaziamento experimental das glândulas das aranhas-criadas-em-tubos (puxando-se os fios diretamente das fiandeiras), por duas vezes, previamente à soltura, eliminava algumas destas diferenças, talvez por ter constituído um estímulo para um aumento da produção de seda. Mesmo assim, permaneciam irregularidades nas teias destas aranhas, que as distinguiam das teias normais, um ponto a favor da hipótese da existência de efeitos de treino. Não sei qual a natureza da seda que Witt, Reed e Scarboro (1969) extraíram das aranhas: provavelmente fosse a das glândulas ampuliformes, usada nas partes secas da teia, não a seda viscosa produzida nas glândulas agregadas e flageliformes (Kovoor, 1977; Kavanagh & Tillinghast, 1979). Se este for o caso, ainda caberia entender a irregularidade no padrão da teia, em aranhas privadas, como dependente da redução de produção em certas glândulas de seda.
Outra manipulação experimental, na mesma pesquisa, consistia em alimentar aranhas na boca, deixando outras usarem sua própria teia para caçar. Trouxe um resultado surpreendente por discrepar dos encontrados por Reed et al. (1970): as aranhas privadas de caçar tinham sua taxa de construção rebaixada em relação à taxa das outras. Era como se a oportunidade de capturar presas na teia tivesse efeitos reforçadores.
Os experimentos de privação feitos até o momento indicam uma notável resistência do algoritmo de construção a condições prévias desfavoráveis para o "treino". Mas também nos põem diante de indícios de plasticidade que merecem ulterior averiguação experimental.
Escorada em diversas constatações - construção precoce, ausência de efeitos da privação drástica (principlamente a partir dos resultados de Petrusewiczowa e de Mayer), caráter típico-da-espécie da teia - a interpretação instintivista da construção da teia é, hoje, predominante. Encontra-se em Foelix (1982), que, depois de colocar a questão "É a construção da teia determinada geneticamente ou é aprendida ? "e depois de considerar que mudanças na teia, ou são produto de uma flexibilidade embutida no programa de construção ou decorrem de processos maturacionais, conclui que "a aprendizagem não parece estar envolvida na (construção) da teia orbicular 'adulta'" (p.143). O próprio Foelix, em outro momento da discussão, espanta-se com o ajustamento do comportamento de construção de A. diadematus às condições de imponderabilidade (experimentos da missão Skylab, Witt, Scarboro, Peakall & Gause, 1977), mas não o toma como indício de capacidade de aprender.
Vollrath (1992) assume uma posição semelhante:
As regras de construção da teia da aranha orbitela foram fixadas em seu genoma por seleção prévia (...) Privando aranhas da experiência de construir de modo algum afeta seu desempenho (...) o que indica que a aprendizagem assume um papel muito pequeno, se é que o assume, na construção da teia (p.148).
Parece-me prematuro, apesar destas opiniões, fechar a questão da plasticidade do comportamento de construção. Os argumentos clássicos - desempenho precoce, experimentos de privação etc. - embora comprovem a contribuição da informação genética para a organização de um desempenho complexo como é a construção da teia - não podem ser tomados como afastando a possibilidade de qualquer efeito de experiência. Mostram não ser relevante uma determinada condição ontogenética.
Idéias orientadoras (Ades, 1987, 1993), contrárias a uma visão dicotômica do comportamento típico-da-espécie, do tipo either-genes-or-environment têm me incentivado, ao longo dos anos, a procurar por fatores de memória e aprendizagem na construção da teia geométrica e em outros aspectos do comportamento de aranhas. A concepção da construção como comportamento orientado, como uma espécie de busca da presa (portanto, como comportamento apetitivo), a flexibilidade e a escolha que parecem reger a fase de escolha do local para construir (Vollrath, 1992), a possibilidade de que a memória controle a escolha dos pontos de colagem da espiral provisória (Eberhard, 1988b), a existência de processos de memória em outros setores do comportamento de aranhas orbitelas (Ades, 1982a, 1982b, 1991; Le Guelte, 1969; Sebrier, Pasquet & Leborgne, 1994a, 1994b), são elementos que justificam mais explorações no campo intrigante da plasticidade comportamental.
Na presente pesquisa, planejei proporcionar a aranhas orbitelas adultas uma experiência diferencial que não envolvesse privação total das rotinas motoras de construção da teia, nem uma restrição drástica à produção de seda pelas glândulas sericígenas. A grupos de aranhas A. argentata seria dado - por um prazo de tempo razoável - contato com ambientes de construção de tamanhos diversos, verificando-se, depois, como constróem, no mesmo ambiente-teste. Resultados de meu laboratório indicam que A. argentata é capaz de ajustar sua teia a caixas-suportes diferentes; que ela é até capaz de distorcer, se premida por condições especiais, algumas das proporções normais de suas teias, produzindo teias oblongas no sentido horizontal ou no vertical (Ades & Rossetto Jr., 1983; Ades, 1986). O espaço disponível para a construção afeta tanto o tamanho das teias como aspectos da estrutura interna (como a excentricidade do miolo, isto é, o quanto se afasta do centro geométrico da teia).
Existem indícios de que experiências de relativo curto-prazo, como a que se pensou usar aqui, possam modificar o desempenho de aranhas adultas. Testando a idéia de que as transformações das teias que surgem com a idade têm a ver com mudanças no peso corporal das aranhas, e retomando um procedimento de Mayer (1953), Christiansen, Baum e Witt (1962), aumentaram em 30 % o peso de espécimes de A. diadematus e A. sericatus, colando-lhes pedaços de chumbo no abdomen. Verificaram que as teias passavam a ter, em relação ao normal, um número menor de raios e um espaçamento maior das espiras. O dispêndio global de seda talvez permanecesse o mesmo, uma vez que os fios pareciam aumentar de grossura, ao mesmo tempo em que perdiam em comprimento. O interessante é que, retirados os pesos, as aranhas não voltaram de imediato a tecer teias dentro do padrão normal: a volta era gradual, como se estivesse envolvido um processo de aprendizagem.
Na fase de aquisição de experiência, no presente experimento, as aranhas A. argentata dos diversos grupos estariam sendo levadas (1) a construir teias diferentes em tamanho e aspectos estruturais, e (2) a gastar quantidades totais diferentes de seda seca e seda viscosa, talvez regulando sua produção diária de seda em função da demanda diferencial. Tanto o desempenho prévio como a regulação da produção de seda poderiam afetar o desempenho na fase de teste, quando as aranhas estivessem, no teste, em caixas-suportes de mesmas dimensões. Poder-se-ia esperar, se houvesse influência da experiência passada, que aranhas treinadas em caixas maiores ocupassem uma parte maior da caixa-suporte de teste do que aranhas treinadas em caixas menores, ou que apertassem mais as voltas de suas espiras viscosas no sentido de fazer uso mais farto da seda. Poder-se-ia também esperar diferenças em características estruturais internas das teias, a partir do desempenho diferente, durante o treino.
Método
Trabalhamos com um lote de 33 aranhas fêmeas adultas (Argiope argentata, Fabricius), capturadas no campus da Universidade ou em bairros próximos da cidade de São Paulo. As aranhas eram colocadas em caixas-suportes de tamanhos diferentes constituídas de um quadro de madeira delimitado, de um lado e de outro, por placas de vidro. Não conseguindo se locomover no vidro, as aranhas acabavam prendendo amarras na madeira e tecendo teias num plano próximo da vertical. Elas recebiam uma mosca por dia, cinco dias por semana, e eram submetidas a um regime de 12 horas de luz/ 12 horas de escuridão. Nesta rotina de laboratório, à qual se adaptam bem as aranhas, teias regulares são construídas a cada 5 dias em média (intervalo: 1 a 11 dias).
As caixas, todas quadradas, com uma profundidade de 8 cm, eram de cinco tamanhos: (1) 12,5 x 12, 5 (área : 156,25 cm2); (2) 17,7 x 17,7 (área 312,5 cm2); (3) 25,0 x 25,0 cm (área: 625 cm2); (4) 35,4 x 35, 4 cm (área: 1250 cm2) e (5) 50 x 50 cm (área: 2500 cm2). Nota-se que a caixa 2 tem o dobro de superfície da caixa 1, que a caixa 3 tem o dobro de superfície da caixa 2 e assim por diante. Resultados de nosso laboratório indicam que espécimes de A. argentata tecem teias regulares nas diversas caixas, aumentando a extensão da teia à medida que cresce o espaço disponível, mudando os índices de excentricidade do miolo e o intervalo inter-espiras.
O procedimento foi implementado em duas etapas. Na fase de experiência prévia, que durou dois meses, cada grupo de aranhas foi exposto a um tamanho de caixa (1, 2, 3, 4 ou 5). Durante esta fase, em que se pretendia que fosse adquirida uma informação a respeito do espaço de construção, as aranhas teceram de 12 a 20 teias. Não sendo as teias destruídas pelos experimentadores, as aranhas podiam utilizar a estratégia de reconstrução da teia, em que cada teia prévia é aproveitada como andaime para a construção da teia ulterior.
Na fase de transferência, as aranhas eram todas colocadas em caixas 3, previamente desembaraçadas de quaisquer teias ou fios. Tínhamos, assim, 5 condições experimentais: 1-3 (n = 6), 2-3 (n = 6), 3-3 (n=7), 4-3 (n=8) e 5-3 (n=6). Nas condições 1-3 e 2-3, as aranhas passavam de uma caixa menor para uma caixa maior; nas condições 4-3 e 5-3, de uma caixa maior para uma caixa menor. A condição 3-3, em que as aranhas eram transferidas de um ambiente para outro de mesmas dimensões, servia de controle.
A última teia da fase de experiência prévia e a primeira teia da fase de transferência foram registradas fotograficamente, usando para isso um equipamento através do qual os fios de seda ganhassem luminosidade máxima e contrastassem com um fundo preto, tornando desnecessário o uso de pó ou tinta para aumentar sua visibilidade (Langer & Eberhard, 1969; Witt, 1971b). Do equipamento constavam (1) uma caixa-fundo de 60 x 60 x 80 cm de profundidade forrada com feltro preto, (2) oito lâmpadas fluorescentes Osram (15 W) dispostas duas a duas na frente e nas bordas da caixa-fundo, formando uma fonte quadrada de raios luminosos e (3) um anteparo dianteiro. A teia da aranha (da qual tinha sido retirada a residente, através de um gentil tapinha no abdomen que provocava queda precipitada num vidrinho de captura) era disposta entre a fonte de luz e o anteparo dianteiro e as fotos eram tiradas com uma câmera Asahi-Pentax e filmes Kodak Panasonic, usando a velocidade indicada pelo sensor automático. De cada foto constavam: (1) um mostrador com data e número de identificação da aranha individual ; (2) um padrão, constituído de dois fios verticais a um centímetro de distância um do outro, que servia de base para uma avaliação das dimensões reais das teias.
A análise das teias foi efetuada a partir de ampliações de aproximadamente 15,5 x 15,5 cm feitas em papel fotográfico de alto contraste. Na foto eram traçados dois eixos retos, um deles vertical (delimitando as partes esquerda e direita da teia), o outro horizontal (delimitando as parte de cima e de baixo da teia), com intersecção no centro do miolo. As medidas seguintes foram obtidas, seja sobre um, seja sobre outro eixo (Figura 1):
Diâmetros vertical e horizontal da teia: distância entre (1) os fios de quadro superior e inferior, (2) os fios de quadro à esquerda e à direita. O diâmetro médio da teia - média de (1) e (2) - foi usado como estimativa do tamanho global da teia.
Diâmetros vertical e horizontal da região da espiral seca: distância entre as espiras secas limítrofes (mais afastadas do miolo) (1) na parte de cima e na parte de baixo da teia, (2) à esquerda e à direita.
Número de espiras viscosas na parte de cima e na de baixo: número de espiras (1) na parte de cima, (2) na parte de baixo da teia. A soma de (1) e (2) foi usada como estimativa do número total de espiras da teia (número de espiras da teia).
Largura da espiral viscosa na parte de cima na de baixo: distância entre a espira viscosa mais distante e a mais próxima do miolo (1) na parte de cima, (2) na parte de baixo da teia.
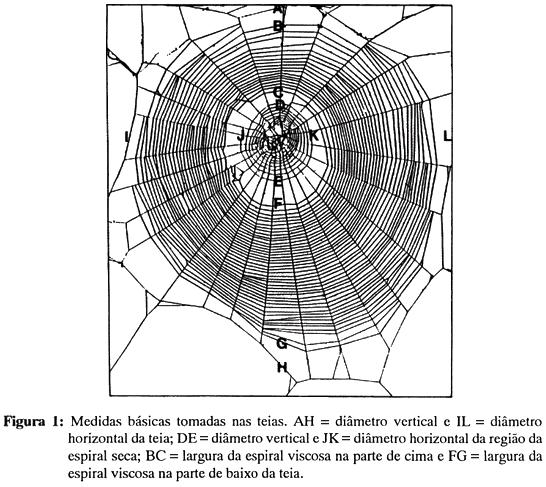
Número de raios.
Calculou-se também a excentricidade vertical da teia (quociente entre a largura da espiral viscosa na parte de cima e na de baixo da teia) e o intervalo inter-espiras (quociente entre a largura da espiral viscosa e o número correspondente de espiras viscosas menos um, na parte de cima e na de baixo da teia). A excentricidade vertical fornece uma indicação de quanto o miolo está situado acima ou abaixo do centro geométrico da teia. O intervalo inter-espira é uma estimativa da densidade dos fios viscosos.
A comparação entre medidas da teia para as diversas condições (1-3, 2-3, 3-3, 4-3 e 5-3) na fase de transferência, meio para testar a hipótese de uma influência da experiência passada, foi efetuada através de uma análise de variância para dados independentes. Para a análise das "regras estruturais" da teia em caixa 3 (fase de transferência) utilizaram-se testes de t de Student para medidas dependentes e correlações de Pearson.
Resultados e discussão
O ajustamento da teia ao espaço disponível
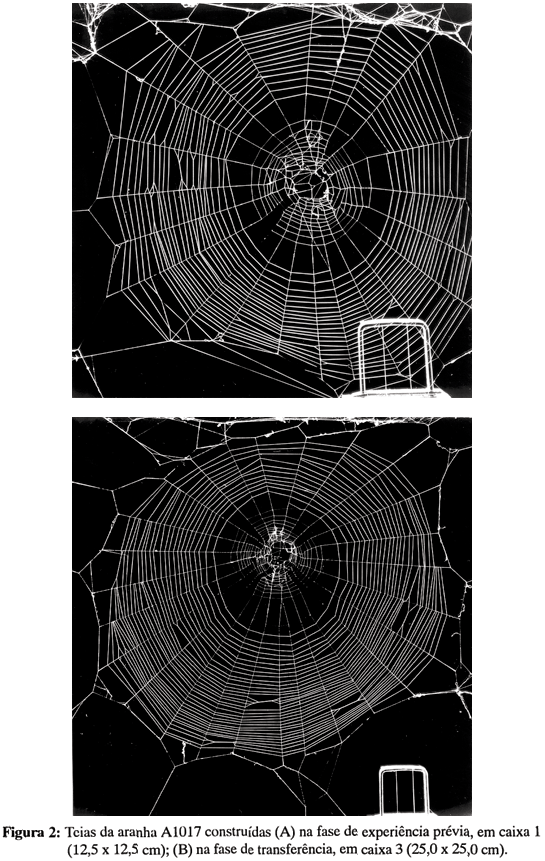
Na fase de experiência prévia, as aranhas construíram teias ajustadas ao tamanho da caixa-suporte, dentro do que se esperaria a partir da pesquisa prévia (Ades, 1986a). A passagem para a caixa 3, na fase de transferência, manifestou-se - exceto para os animais que vinham de uma caixa de tamanho semelhante - por uma mudança marcada nas dimensões e no padrão das teias. As figuras 2 e 3 mostram dois casos representativos. São fotos das teias de duas aranhas, uma da condição 2-3, outro de uma aranha na condição 4-3. Levando em conta o padrão de 1 cm para efetuar as comparações, vê-se que a aranha A1017 utilizou um comprimento maior de fios, tanto secos como viscosos, na feitura da teia em caixa 3 do que em caixa 2, e que a aranha A1021, ao contrário, diminuiu o comprimento de fio utilizado, quando passou da caixa 4 para a caixa 3. Estes resultados também sugerem que a quantidade de seda gasta tenha diferido, de uma caixa para outra, em ambos os animais.

Experimentos vários indicam que a aranha construtora está sob o controle da quantidade de seda de que dispõe, no início da construção. Eberhard (1988a), trabalhando na Costa Rica com Leucauge mariana, uma aranha que pode ter dois, até três episódios de construção no mesmo dia, cortava a maior parte dos raios, em teias recém - elaboradas, forçando a aranha a refazer sua rede usando o mesmo arcabouço. As teias de substituição eram significativamente menores, tinham menor número de raios e de espiras viscosas, do que as teias iniciais. Terceiras teias (se o espírito científico levasse o experimentador a também arrombar as segundas) eram menores ainda. Se as teias iniciais fossem destruídas antes que a aranha tivesse tido tempo de colocar a espiral viscosa (isto é, deixando intacto o suprimento das glândulas agregadas e flageliformes, produtoras de fio viscoso) o prejuízo era menor, a aranha mantinha, nas teias de substituição, o tamanho de suas teias normais. Claramente, orbitelas "decidem" as dimensões de suas teias em função da quantidade de seda disponível, a partir de um provável feedback sobre o estado de preenchimento das glândulas envolvidas.
Os presentes resultados sugerem a existência de um critério complementar: as aranhas regulam a quantidade de seda a ser gasta a partir dos parâmetros ambientais. Ao terminar a feitura de sua teia em caixa 1, uma Argiope permanece provavelmente com um excedente de seda nas fiandeiras maior do que após a construção em caixa 3. Somente em ambientes bastante grandes, como em caixas 5, é que a quantidade de seda disponível estabelece um limite para a ocupação do espaço: a teia atinge seu máximo de extensão.
Ausência de efeitos da experiência passada
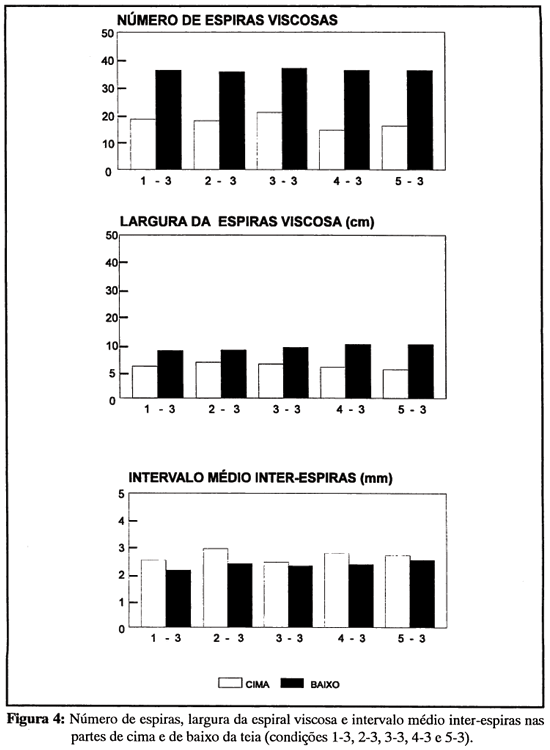
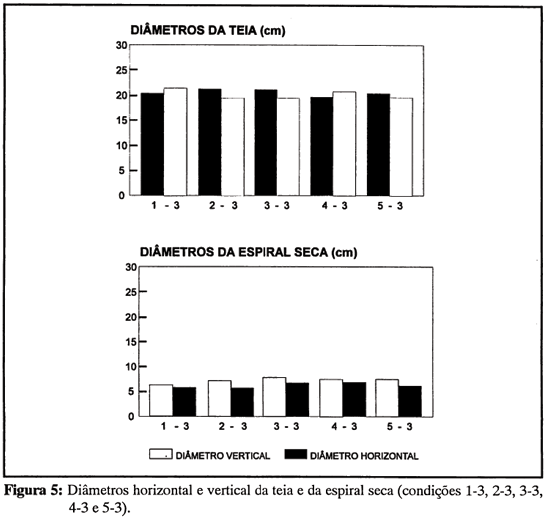
Não houve indício algum de que a experiência de construção em caixas de tamanhos diferentes (1, 2, 3, 4 e 5) tivesse influenciado, na fase de transferência, a feitura da teia em caixa 3. Ao contrário, impressiona a homogeneidade das medidas da teias em caixa 3, de uma condição experimental para outra (figuras 4 e 5). Todas as aranhas - com exceção, obviamente, das do grupo 3-3 - modificaram o tamanho de sua teia quando, na passagem da fase de experiência prévia para a de transferência, foram forçadas a mudar de ambiente. Mas a modificação, seja para menos (condições 4-3 e 5-3), seja para mais (condições 1-3 e 2-3) redundou em teias semelhantes. Não foram encontradas diferenças estatísticas significativas, entre condições, no diâmetro vertical (F4,28 = 0,31, p = 0,85) ou horizontal da teia (F4,28 = 0,63, p = 0,65), no diâmetro vertical (F4,28 = 0,55, p = 0,70) ou horizontal da região da espiral seca (F4,28 = 0,34, p = 0,82), no número de espiras viscosas na parte de cima (F4,28 = 0,41, p = 0,80) ou de baixo da teia (F4,28 = 0,08, p = 0,99), na largura da espiral viscosa na parte de cima (F4,28 = 0,36, 0 = 0,83) ou de baixo da teia (F4,28 = 0,70, p = 0,60), e no número de raios (F4,28 = 0,37, p = 0,83) das teias feitas em caixa 3.
Deve-se concluir que, para a aranha construtora, pelo menos nas condições de nosso estudo, são as dimensões presentes do espaço disponível que contam. É em relação a estas dimensões - avaliadas, de uma forma ou de outra, através de idas e vindas exploratórias - que a aranha calibra os parâmetros básicos de sua teia, mantendo inalterado o algoritmo comportamental.
Regras de construção em caixas 3
Uma vez que basicamente semelhantes entre condições (1-3, 2-3, 3-3, 4-3 e 5-3), as teias em caixa 3 da fase de transferência puderam ser tomadas em conjunto e submetidas a análise para determinar algumas das relações estruturais ou regras de construção da teia (Peters, 1954). Estas regras representam a maneira como a aranha utiliza o espaço disponível e, especialmente, a maneira como dispõe e modula reciprocamente as diversas partes de sua rede.
Regras de tamanho e quantidade. Algumas das regras de construção têm a ver com tamanho (da teia ou de suas partes) ou com quantidade de elementos. A teia, ou suas regiões, tendem a aumentar proporcionalmente no sentido vertical e horizontal. Encontra-se, nas caixas 3, uma correlação positiva significativa entre o diâmetro vertical da teia e o seu diâmetro horizontal (r = 0,558, p < 0,01) e outra entre o diâmetro vertical da região da espiral seca e o seu diâmetro horizontal (r = 0,626, p < 0,01).
O tamanho global da teia, estimado através da média entre os diâmetros vertical e horizontal, correlaciona-se positivamente com diversas medidas da teia, o que sugere que a colocação dos primeiros fios do arcabouço seco da teia, fios de quadro, raios e amarras, determina um espaço de trabalho que serve de guia ou restrição para as etapas subseqüentes de construção.
Este efeito de regulação das partes pela configuração do arcabouço se verifica para os raios, para a espiral seca, para a espiral viscosa. Quanto maior o valor do diâmetro médio da teia, (1) maior o número de raios (diâmetro médio da teia x número de raios, r = 0,357, p < 0,05); (2) maior o tamanho da região da espiral seca (diâmetro médio da teia x diâmetro médio da região da espiral seca, r = 0, 384, p < 0,05); (3) maior o número de espiras viscosas (diâmetro médio da teia x número de espiras viscosas, r = 0552, p < 0,01). Correlações positivas significativas também são encontradas entre o diâmetro médio e as larguras da espiral viscosa. Nota-se que os valores de r não são altos: outros fatores, além da extensão do arcabouço básico da teia, provavelmente intervêm nas decisões quantitativas da construção.
Resultados semelhantes foram obtidos, entre outros pesquisadores, por Tilquin (1942) em teias de Argiope bruennichi e Araneus quadratus; por Peters (1954) em teias de Micrathena gracilis, Gasteracantha cancriformis, Cyclosa trifida, Cyclosa caroli e Argiope argentata; por Eberhard (1972) em teias Uloborus diversus.
Regras de assimetria vertical. As teias orbiculares contruídas em plano vertical ou próximo dele, como as de A. argentata, marcam-se pela assimetria vertical: a parte de cima (sobre o miolo) é geralmente menor do que a da baixo. Esta assimetria se verifica, nas caixas 3, para as medidas da espiral viscosa (Figura 4). A largura em cima desta espiral difere significativamente da largura em baixo (t = 8,38, gl = 32, p < 0,0001). O número de espiras em cima difere significativamente do número de espiras em baixo (t = 13,12, gl = 32, p < 0,0001).
Outra assimetria tem a ver com o espaçamento médio das espiras viscosas (Figura 4). Em cima, as espiras distam, em média, 2,5 mm; em baixo, 2,1 mm. Argiope argentata parece não estar controlada pelo mesmo conjunto de variáveis, quando em cima, quando em baixo da teia, na fase de fixação das espiras viscosas. Esta suposição recebe apoio da análise correlacional: a correlação entre a largura da espiral viscosa na parte de cima e o número de espiras em cima (r = 0,863, p < 0,01) é significativa; a da parte de baixo, não (r = 0,2515, p > 0,05). Os espaçamentos, na parte de cima, são menos variáveis do que os da parte de baixo. Fatores como a quantidade de fio disponível, a excentricidade da teia ou outros poderiam estar atuando, preferencialmente, na parte de baixo da teia, introduzindo variação. Assimetrias cima/baixo na densidade de fios viscosos também foram notadas, em Araneus diadematus, por Vollrath (1992).
O caráter elítico da teia ou de suas estruturas (do mesmo modo que as assimetrias verticais) significa um provável controle da construção pela gravidade. Se este caráter elítico é verificado na região da espiral seca, cujo diâmetro vertical é maior que o horizontal (t = 7,081, gl = 32, p < 0,0001), não o é na teia como um todo (Figura 5). Os diâmetros vertical e horizontal, com aproximadamente 21 cm de comprimento, não diferem significativamente (t = 0,060, gl = 32, p > 0,05).
A construção da teia enquanto instinto?
Os resultados do presente experimento causaram uma certa frustração a quem esperava demonstrar plasticidade na construção da teia geométrica. A quase identidade das teias feitas por aranhas de experiência prévia diferente depõe fortemente contra a hipótese de que esta experiência tenha algum papel no desempenho e reforça a hipótese de que o cálculo espacial para a colocação dos fios seja feito a partir do contexto imediato dos suportes.
Isto não significa que processos de memória não estejam mobilizados, durante a construção. Ao explorar uma caixa nova, através de idas e vindas, de subidas e descidas, em movimentos de aparente desordem, A. argentata provavelmente adquira informações sobre o espaço que lhe é disponível e modifique, de modo a ajustá-lo, seu comportamento de tecer os primeiros fios do arcabouço da teia. As perguntas de Vollrath (1992) a respeito da fase exploratória da construção ("Será que a informação espacial é levada em conta? Se de fato é levada em conta, como é conseguida, em que nível de pormenor? De que maneira é ela armazenada na memória da aranha? Seria possível que fosse na forma de um mapa espacial?"), assim como os resultados de Eberhard (1988b) sobre as trajetórias de L. mariana durante a feitura da espiral provisória, levantam a possibilidade de existir um nível de funcionamento cognitivo em aranhas construtoras. Observações sobre a interrupção e retomada do construir também falam a favor da adoção de hipóteses mais sofisticadas a respeito de como se integra o comportamento das aranhas orbitelas (Ades, 1982b).
Mas a memória eventualmente usada permanece presa ao desempenho imediato, sua influência não é transferida às posteriores feituras de teias. Está como que embutida no algoritmo de construção e zera-se com o término das estruturas em que seu efeito se manifesta. Participa do ajustamento da teia ao contexto presente de suportes e fios mas não gera um pré-cálculo de dimensões e relações estruturais que se esperaria atuassem, se verdadeira a hipótese de aprendizagem. Funciona como memória operacional, não como memória de referência (Xavier, 1993).
Quer incorpore mecanismos de memória ou não, a construção da teia parece resistir à influência de certos contextos ontogenéticos. Destaquei a palavra "certos" pois carrega duas idéias relevantes do ponto de vista da velha dicotomia inato/aprendido. Há toda uma epistemologia a ser extraída de experimentos sobre teias de aranhas. De um lado, a palavra certos significa incerteza: remete ao argumento de incompletude de qualquer experimento que mantenha o organismo privado de experiência ou que manipule (sem privação drástica, como foi o caso no presente estudo) sua história de desempenhos e contatos com o ambiente. Sempre se poderá arguir que esta filtragem da experiência não tenha sido suficiente, que outros fatores de experiência estavam ou poderiam estar envolvidos na ontogênese do desempenho concreto. Até aqui, estou com os teóricos que reagem ao que há de absoluto nas formulações clássicas (e nas acepções do senso comum) acerca de comportamento instintivo.
A palavra transmite, de outro lado, uma verdade ontogenética: que os eventos passados não são todos aptos a modificar o desempenho de forma plástica, e que alguns certamente não o modificam. Mostrando que um roteiro de experiência passada não altera - em relação a outro roteiro, utilizado como critério - um desempenho, não autoriza a dizer que o desempenho, ou os processos que lhe subjazem, é um instinto. Mas permite afirmar que é independente da experiência, no que tange às variáveis manipuladas. A rejeição do pensamento either-genes-or-environment ou de qualquer outra dicotomização não significa que não seja possível realizar uma análise rigorosa da ontogênese, distinguindo os fatores aos quais o desempenho é e não é sensível.
A aceitação deste argumento elimina a contradição que parece existir na afirmação de que dado desempenho é, ao mesmo tempo, plástico e não-plástico ou, para usar uma linguagem há tanto tempo em vias de se tornar obsoleta, instintivo e capaz de sofrer efeitos de experiência. Proponho que se troquem proposições relativas ao desempenho como um todo por proposições contextualizadas, que tenham a ver com aspectos dos processos que o desempenho incorpora.
Os resultados do presente experimento, tomo-os como base empírica para a primeira parte de meu argumento: a construção da teia, em aranhas orbitelas, não é plástica em certos aspectos de sua produção, o ajustamento espacial que ela expressa depende de uma leitura do ambiente imediato.
Para que o argumento se feche, é necessário passar à contra-proposição: a construção da teia, em aranhas orbitelas, pode depender de experiência, no bom e velho sentido de uma aquisição relativamente duradoura de informação (capaz, por exemplo, de transferir-se de um episódio de construção a outro). Os primeiros resultados de um programa de pesquisa sobre o ajustamento de Argiope argentata a ambientes gravitacionais modificados (Ades & Cunha, 1991; Ades, Cunha & Tiedemann, 1993) parecem indicar que, sim, há efeitos de experiência na construção da teia. Esta segunda parte do argumento, deixo para outro artigo.
Nota:
1. O artigo é dedicado a Walter Hugo de Andrade Cunha, orientador de minha tese de doutorado sobre Argiope argentata, colega e amigo. Guardo ainda as versões dos capítulos desta tese, com extensas anotações em vermelho no verso de cada página (não caberiam na margem) todas pertinentes e instigantes. Muitos eram os pontos em que Walter indicava, com delicadeza, sugestões de mudança do texto ou discordâncias ou dúvidas : "não me parece uma boa justificação...", "penso que seria melhor dizer...", "parece contraditório com o dito acima", "e quanto à variação individual, não há ?", etc. Elogios vinham no momento certo, destacando aspectos dotados de implicações teóricas ou dados surpreendentes. "Tudo muito bonito (até poético) !" anotava Walter, numa das páginas-ao-lado da versão do capítulo sobre teias. E, no final do mesmo : "Está excepcionalmente rica e bem desenvolvida esta parte do trabalho, além de magnificamente discutida". Serei mais do que gratificado se o presente estudo merecer dele a metade de uma opinião tão generosa.
REFERÊNCIAS BIBLIOGRÁFICAS
ADES, C. A construção da teia geométrica como programa comportamental. Ciência e Cultura, v.38, p.760-75, 1986. [ Links ]
ADES, C. Immediate stimulation and recent experience as controlling factors in the predatory sequence of the spider Argiope argentata (Fabricius). Interamerican Journal of Psychology, v.16, p.57-63, 1982a. [ Links ]
ADES, C. Interrupção e retomada da construção da teia numa aranha orbitela: um estudo descritivo. Ciência e Cultura, v.34, n.7, p.900, 1982b. Suplemento. /Apresentado à 34. Reunião Anual da SBPC, Campinas, 1982 - Resumo/ [ Links ]
ADES, C. Memória e instinto no comportamento de predação da aranha Argiope argentata. São Paulo, 1991. 132p. Tese (Livre Docência) - Instituto de Psicologia, Universidade de São Paulo. [ Links ]
ADES, C. Notas para uma análise psicoetológica da aprendizagem. Boletim de Psicologia, v.37, n.86, p.24-35, 1987. [ Links ]
ADES, C. Por uma história natural da memória. Psicologia USP, v.4, n.1/2, p.25-47, 1993. [ Links ]
ADES, C. A teia e a caça da aranha Argiope argentata. São Paulo, 1972. 493p. 2.v. Tese (Doutorado) - Instituto de Psicologia, Universidade de São Paulo. [ Links ]
ADES, C.; CUNHA, S.S. O ajustamento da teia da aranha Argiope argentata a condições gravitacionais modificadas. In: ENCONTRO ANUAL DE ETOLOGIA, 8., Natal, Rio Grande do Norte, 1991. Anais de etologia. Natal, 1991. p.24-6. [ Links ]
ADES, C.; CUNHA, S.S.; TIEDEMANN, K. Experience-induced changes in orb-web building. In: INTERNATIONAL CONGRESS OF ETHOLOGY, 23., Torremolinos, Espanha, 1993. Abstracts. Torremolinos, 1993. p.122. [ Links ]
ADES, C.; ROSSETTO Jr., J.A. Espaço e programa de construção da teia numa aranha orbitela: resultados preliminares. In: ENCONTRO PAULISTA DE ETOLOGIA, 1., Jaboticabal, São Paulo, 1983. Anais de Etologia. Jaboticabal, 1983. p.247. [ Links ]
ADES, C.; ROSSETTO Jr., J.A. Pode a experiência passada a curto prazo influir na construção da teia geométrica? Ciência e Cultura, v.36, n.7, p.974, 1984. Suplemento. /Apresentado à 36. Reunião Anual da SBPC, São Paulo, 1984 - Resumo/ [ Links ]
BERLAND, J. Adaptation de l'instinct chez une Araignée: Nemoscolus laurae E. Simon. Archives de Zoologie Expérimentale, v.56, p.134-8, 1917. [ Links ]
CHRISTIANSEN, A.; BAUM, R.; WITT, P.N. Changes in spider webs brought about by mescaline, psilocybin and an increase in body weight. Journal of Pharmacology and Experimental Therapy, v.136, p.31-7, 1962. [ Links ]
CRAIG, C.L. Predator foraging behavior in response to perception and learning by its prey: interactions between orb-spinning spiders and stingless bees. Behavioral Ecology and Sociobiology, v.35, p.45-52, 1994. [ Links ]
CRAIG, C.L.; BERNARD, G.D. Insect attraction to ultraviolet-reflecting spiders and web decorations. Ecology, v.71, p.611-23, 1990. [ Links ]
EBERHARD, W.G. Behavioral characters for the higher classification of orb-weaving spiders. Evolution, v.36, p.1067-95, 1982. [ Links ]
EBERHARD, W.G. Behavioral flexibility in orb web construction: effects of supplies in different silk glands and spider size and weight. Journal of Arachnology, v.16, p.295-302, 1988a. [ Links ]
EBERHARD, W.G. Function and phylogeny of spider webs. Annual Review of Ecology and Systematics, v.21, p.341-72, 1990. [ Links ]
EBERHARD, W.G. Memory of distances and directions moved as cues during temporary spiral construction by the spider Leucauge mariana (Araneae, Araneidae). Journal of Insect Behavior, v.1, p.51-66, 1988b. [ Links ]
EBERHARD, W.G. The web of Uloborus diversus (Araneae: Uloboridae). Journal of Zoology, v.166, p.417-65, 1972. [ Links ]
FERNANDES, M. Hai-Kais. São Paulo, Ed. Senzala, s.d. [ Links ]
FOELIX, R.F. Biology of spiders. Cambridge, Harvard University Press, 1982. [ Links ]
KASTON, B.J. The evolution of spider webs. American Zoologist, v.4, p.191-207, 1964. [ Links ]
KAVANAGH, E.; TILLINGHAST, E.K. Fibrous and adhesive components of the orb webs of Araneus trifolium and Argiope trifasciata. Journal of Morphology, v.160, n.1, p.17-25, 1979. [ Links ]
KOVOOR, J. La soie et les glandes séricigènes des arachnides. Année Biologique, v.16, p.97-171, 1977. [ Links ]
LAHUE, R. The chelicerates. In: CORNING, W.C.; DYAL, J.A.; WILLOWS, A.O.D. Invertebrate learning. New York, Plenum Press, 1973. v.2: Arthropods and gasropod mollusks. [ Links ]
LANGER, R.M.; EBERHARD, W. Laboratory photography of spider silk. American Zoologist, v.9, p.97-101, 1969. [ Links ]
LE GUELTE, L. Learning in spiders. American Zoologist, v.9, p.142-52, 1969. [ Links ]
LE GUELTE, L. Sur le comportement des Araignées libérées du cocon avant la date normale. Comptes Rendus de l'Académie des Sciences, v.266, p.382-3, 1968. [ Links ]
LEVI, H.W. Orb-weaving spiders and their webs. American Scientist, v.66, n.6, p.734-42, 1978. [ Links ]
MAYER, G. Untersuchungen uber Herstellung und Struktur des Radnetzes von Aranea diadema und Zilla x-notata mit besonderer Berucksichtigung des Unterschied von Jugend- und Altersnetzen. Zeitschrift fur Tierpsychologie, v.9, p.337-62, 1953. [ Links ]
PETERS, H. Contribuciones sobre la etología y ecología comparada de las arañas tejedoras tropicales. Comunicaciones del Instituto Tropical de Investigaciones Científicas (El Salvador), v.4, p.37-46, 1955. [ Links ]
PETERS, H. Estudios adicionales sobre la estructura de la red concentrica de las arañas. Comunicaciones del Instituto Tropical de Investigaciones Científicas (El Salvador), v.3, p.1-18, 1954. [ Links ]
PETERS, H. Maturing and coordination of web-building activity. American Zoologist, v.9, p.223-7, 1969. [ Links ]
PETERS, H. Studien an Metz der Kreuzspinne (Aranea diadema). Zeitschrift fur Morphologie und Ökologie der Tiere, v.32, p.613-49, 1936. [ Links ]
PETERS, H. Untersuchungen ueber die Proportioniergun im Spinnen-Netz. Zeitschrift fur Naturforschung, 1954. [ Links ]
PETRUSEWICZOWA, E. Beobachtungen uber den Bau des Netzes der Kreuzspinne. Travaux de l'Institut de Biologie de l'Université de Wilno, v.9, p.1-25, 1938. [ Links ]
PORTER, J.P. The habits, instincts and mental powers of spiders, genera Argiope and Espeira. American Journal of Psychology, v.17, p.306-57, 1906. [ Links ]
RAMOUSSE, R. Le comportement constructeur chez les araignées orbitèles. Journal de Psychologie Normale et Pathologique, n.3, p.217-39, 1982. [ Links ]
RAMOUSSE, R.; WURDAK, E. Biologie et comportement de la larve et de la première nymphe de l'araignée Araneus suspicax (O. Pickard-Cambridge). Revue Arachnologique, v.5, p.275-86, 1984. [ Links ]
REED, C.F.; WITT, P.N.; SCARBORO, M.B.; PEAKALL, D.B. Experience and the orb-web. Developmental Psychobiology, v.3, n.4, p.251-65, 1970. [ Links ]
SEBRIER, M.A.; PASQUET, A.; LEBORGNE, R. Resource management in the orb-weaving spider Zygiella x-notata. I. Influence of spider behaviour and available prey. Ethology, v.96, p.334-42, 1994a. [ Links ]
SEBRIER, M.A.; PASQUET, A.; LEBORGNE, R. Resource management in the orb-weaving spider Zygiella x-notata. II. Ability to manage a set of various prey. Ethology, v.96, p.343-52, 1994b. [ Links ]
SHEAR, W.A. Untangling the evolution of the web. American Scientist, v.82, p.256-66, 1994. [ Links ]
TILQUIN, A. La toile géométrique des Araignées. Paris, Presses Universitaires de France, 1942. [ Links ]
VOLLRATH, F. Analysis and interpretation of orb spider exploration and web-building behavior. Advances in the Study of Behavior, v.21, p.147-94, 1992. [ Links ]
VOLLRATH, F. Spiral geometry in the garden spider's orb web. Naturwissenchaften, v.72, p.666-7, 1985. [ Links ]
WITT, P.N. Drugs alter web-building of spiders. Behavioral Science, v.16, p.98-113, 1971a. [ Links ]
WITT, P.N. Instructions for working with web building spiders in the laboratory. BioScience, v.21, p.23-5, 1971b. [ Links ]
WITT, P.N. Specificity of the web. In: WITT, P.N.; REED, C.F.; PEAKALL, D.B., orgs. A spider's web: problems in regulatory biology. New York, Springer Verlag, 1968. [ Links ]
WITT, P.N.; RAWLINGS, J.O.; REED, C.F. Ontogeny of web-building behaviour in two orb-weaving spiders. American Zoologist, v.12, n.3, p.445-54, 1972.
WITT, P.N.; REED, C.F.; PEAKALL, D.B., orgs. A spider's web: problems in regulatory biology. New York, Springer Verlag, 1968. [ Links ]
WITT, P.N.; REED, C.F.; SCARBORO, M.B. Differences in web-patterns between tube- and cage-raised spiders (Araneus diadematus, Cl.). American Zoologist, v.9, p.1061,1969. [ Links ]
WITT, P.N.; SCARBORO, M.B.; PEAKALL, D.B.; GAUSE, R. Spider web-building in outer space: evaluation of records from the Skylab spider experiment. American Journal of Arachnology, v.4, p.115-24, 1977. [ Links ]
XAVIER, G.F. A modularidade da memória e o sistema nervoso. Psicologia USP, v.4, n.1/2, p.61-115, 1993. [ Links ]
ZSCHOKKE, S. The influence of the auxiliary spiral on the capture spiral in Araneus diadematus Clerck (Araneidae). Bulletin of the British Arachnological Society, v.9, p.169-73, 1993. [ Links ]
1 Os dados do presente artigo provêm de pesquisa realizada com José Augusto Rosseto Jr., então estagiário no laboratório de Psicoetologia, a quem agradeço muito. Quero também agradecer a Eduardo Novaes Ramires e a Ana Amélia Benedito Silva pela leitura crítica do texto. O trabalho, que contou com o apoio do CNPq, teve seus dados preliminares apresentados em reunião da SBPC (Ades & Rossetto Jr., 1984).
