










Serviços Personalizados
artigo
 Português (pdf)
Português (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
Indicadores

Compartilhar

 Permalink
PermalinkPsicologia USP
versão On-line ISSN 1678-5177
Psicol. USP v.6 n.1 São Paulo 1995
ARTIGOS ORIGINAIS
Reprodução, alimentação e reatividade em planárias sob diferentes condições de manutenção1
Reproduction, eating and reaction to food in planarians under different maintenance conditions
Inês Amosso Dolci-Palma
Departamento de Psicologia Evolutiva, Social e Escolar - Faculdade de Ciências e Letras de Assis, UNESP
RESUMO
O objetivo do estudo foi analisar alguns efeitos de três diferentes condições de manutenção de planárias em laboratório. Uma das condições era mais estável, padronizada e pobre em possibilidades de estimulação do que a outra. Na terceira condição, as possibilidades de estimulação eram enriquecidas semana a semana. Os grupos de planárias mantidos em cada uma das três condições foram observados durante 35 dias e comparados entre si quanto ao comprimento dos animais, à oviposição e ao modo como as planárias reagiam ao alimento. Na tentativa de compreender as similaridades e as diferenças entre os grupos, foram consideradas implicações contidas em cada uma das formas de manutenção estudadas.
Descritores: Planárias. Análise do comportamento. Reprodução. Alimentação. Desenvolvimento físico. Etologia animal.
ABSTRACT
The purpose of this study was to analyse some effects of three different maintenance conditions of planarians in the laboratory. One of the conditions was stable, standardized and with less possibilities of stimulation than the other. In the third condition the possibilities of stimulation were enriched weekly. The groups of planarians kept in each condition were observed for 35 days and were compared regarding the lenght of the animals, oviposition and reactions to food. The implications of the different forms of maintenance were considered in the discussion of the similarities and differences between the groups.
Index terms: Planarians. Behavioral assessment. Reproduction. Eating. Physical development. Animal ethology.
Os Turbellaria, uma das três classes do filo Platyhelminthes, compreendem vermes de vida livre, habitantes de água doce e salgada e de terras úmidas.
As planárias de água doce, pertencentes a essa classe, habitam águas paradas ou com pouca correnteza como beiras de riachos. São encontradas por entre as ramagens e as raízes da vegetação submersa e, também, sob pedras. Tendem a permanecer nas regiões mais sombreadas do ambiente, visto que são fotonegativas.
Uma vez que se trata de um organismo de pequenas proporções - o verme adulto tem aproximadamente 14 milímetros de comprimento - e que vive em ambientes com as características acima referidas, pode-se depreender que é extremamente difícil o conhecimento etológico dessas planárias em seu próprio habitat.
Na literatura especializada, há muitas informações relativas ao comportamento de planárias. Podemos mencionar trabalhos sobre efeitos de estimulação luminosa (Stewart, 1968; Walter, 1907), sobre formação e manutenção de agregados (Silva, 1976, 1978; Reynierse, Gleason & Otteman, 1969), sobre reatividade a estímulos químicos (Mason, 1975; Pearl, 1903) e a estímulos mecânicos (Bellamy & Reynoldson, 1974; Pearl, 1903). Alguns dos estudos referidos na literatura dizem respeito à aprendizagem instrumental (Corning, 1966; Micklin & May, 1975) e muitos se referem à possibilidade ou não de condicionar planárias segundo o paradigma pavloviano (Jacobson, Horowitz & Fried, 1967; Levison & Gavurin, 1979; Yaremko & Kimmel, 1969). Relacionados a estes últimos, há estudos que investigam as questões polêmicas sobre possibilidade ou não de retenção de uma resposta condicionada (Cherkashin, Sheiman & Bogorovskaya, 1966; McConnell, Jacobson & Kimble, 1959) e de transferência de aprendizagem por canibalismo (Carney & Mitchell, 1978; Loomis & Napoli, 1975). As investigações a que acabamos de nos referir têm sido feitas em situações experimentais criadas em laboratório. Cuidados especiais têm sido tomados nesses estudos quanto ao refinamento dos procedimentos de controle de variáveis relativas quer ao indivíduo, quer ao meio ambiente. Porém, sequer breves referências têm sido feitas às condições em que os animais estudados são mantidos no laboratório.
Considerando a dificuldade em obtermos um referencial consistente oriundo do conhecimento dos modos de ação usuais das planárias em seu habitat e considerando que o cativeiro pode afetar e mesmo deformar o comportamento do animal, julgamos que as condições de manutenção de planárias em laboratório devem ser investigadas de forma mais sistemática do que tem sido feito até agora. Tais condições não podem ser desconsideradas, principalmente nas ocasiões em que o objetivo é determinar os efeitos de situações experimentais sobre o comportamento, pois podem afetá-lo.
Tendo em mente tais preocupações, centramos esta investigação no estudo de algumas características do ambiente de manutenção e em suas relações com o comportamento de planárias, com vistas a auxiliar na decisão sobre como tais animais - que podem vir a ser objeto de experimentos - deveriam ser mantidos em cativeiro, no laboratório.
Como objetivo específico do estudo, propusemo-nos a verificar possíveis efeitos de três diferentes formas de manutenção de planárias do gênero Dugesia em laboratório sobre alguns dos seus comportamentos - relacionados à reprodução e à alimentação - e sobre o seu aspecto físico.
Uma das condições de manutenção aqui empregadas, e denominada de condição de manutenção em grupo, guardava semelhança, pelos seus elementos componentes, com condições tipicamente presentes no habitat das planárias. Durante o transcorrer da investigação, essa condição de manutenção manteve-se relativamente estável e padronizada.
Por oposição a essa condição, criamos a condição de manutenção em isolamento. Tratava-se de uma forma de manter planárias mais pobre, estável e padronizada do que a primeira, quer pela menor diversidade, quer pelo menor número de elementos presentes esporadicamente ou o tempo todo no recipiente onde estavam os animais.
Pensamos que, caso se constatassem diferenças entre esses dois grupos de planárias, caberia perguntar a que elementos presentes ou ausentes em cada uma das duas situações poderiam ser atribuídas tais diferenças.
Decidimos, então, observar um terceiro grupo de planárias, aquelas da condição de manutenção mutável. Esse grupo foi mantido, de início, de modo igual àquele das planárias em isolamento. Gradativamente, a condição de manutenção foi alterada de modo a estar, no final do estudo, em situação igual àquela dos animais da condição em grupo. Portanto, essa condição de manutenção enriquecia-se semana a semana, em termos de possibilidade de estimulação disponível às planárias. A condição de manutenção mutável era, assim, menos estável e padronizada do que as outras duas condições de manutenção empregadas neste estudo.
Com os animais mantidos nesses três ambientes, foram feitas observações do aspecto físico das planárias, da reatividade à alimentação com fígado e da reprodução.
Podemos considerar que a reprodução com sucesso é um indicador de adaptação ao ambiente. Claro está que o conceito de sucesso reprodutivo, tal como o propôs Simpson (1958), não pode ser totalmente aplicado neste trabalho, pois o período de tempo por ele compreendido impede afirmativas acerca de fertilidade dos descendentes, item que deve estar presente para que se possa, com toda propriedade, usar tal conceito. Porém, a duração do estudo permitiu que analisássemos se houve ou não oviposição, qual o seu ritmo e se houve ou não descendentes vivos. Assim, pelo menos parcialmente, pudemos avaliar se a reprodução teve ou não sucesso.
Já com relação à análise de reações de planárias à alimentação com pedaços de fígado cru de boi, embora não dispondo de muitas informações de literatura, tínhamos, tanto uma prática extensa com o uso dessa forma de alimentação, quanto algumas descrições do comportamento de planárias na situação. Esses fatos refletiram-se, no caso, em nossa forma de proceder, ou seja, no fato de termos optado por colocar o fígado nos frascos durante o transcorrer da observação.
Sabíamos que, assim que um pedaço de fígado é colocado na base do recipiente onde estão as planárias, quase que imediatamente elas começam a dirigir-se ao fígado e, em pouco tempo, a maior parte dos animais do frasco está se alimentando. Essas nossas observações concordam com relatos descritivos (Pearl, 1903) a respeito.
Introduzindo o fígado após decorrida metade do tempo da observação, poderíamos comparar o comportamento dos animais antes e depois dessa introdução. Portanto, teríamos condições de considerar, com base em dados agora quantitativos, o quão imediatas seriam as reações das planárias à colocação do alimento na água, como se estenderiam ou se transformariam essas reações ao longo de um certo período de tempo e como essas reações variariam nas diferentes condições de manutenção.
Em laboratório, planárias geralmente são alimentadas com pedaços de invertebrados. A alimentação, duas vezes por semana, com fígado, foi por nós empregada uma vez que é também adequada, conforme dados da literatura (Galtsoff, 1959), para manter animais em laboratório.
Wulzen e Bahrs (1931) investigaram efeitos deletérios relacionados a diferentes balanceamentos da dieta alimentar de planárias. Os efeitos observados foram formação de cistos pelo corpo, podendo levar até mesmo à desintegração do animal, irregularidades nas bordas e escurecimento e enrugamento dos vermes. Da observação desses efeitos, Wulzen e Bahrs concluíram que há um fator, provavelmente uma vitamina, que deve estar presente na dieta do animal que posteriormente terá seu fígado, coração ou rim usado como alimento para a planária, para que esses órgãos se constituam em uma dieta alimentar adequada.
Uma vez que estávamos fornecendo maior quantidade, pelo menos potencialmente, de alimentação aos animais do que aquela normalmente considerada como adequada para manutenção de planárias, interessávamo-nos em observar se isso se refletiria nas condições físicas do animal e, em caso afirmativo, como o faria. Também nos interessava verificar se a condição de manutenção, em geral, afetaria o físico do animal, e, caso o fizesse, como o faria.
Baguna (1974, 1976) demonstrou que Dugesia mediterranea e Dugesia lugubris crescem em função da quantidade e da qualidade da alimentação. Quando o organismo atinge seu tamanho adulto, alimentação e, também, temperatura são fatores importantes para que haja manutenção de um certo tamanho. Baguna analisou ainda a relação entre alimentação e mudanças no índice de mitoses, embora tenha apontado que o processo de aumento do índice mitótico posteriormente à alimentação é ainda desconhecido. O autor supôs a existência de células indiferenciadas no mesênquima, as quais estão prontas para mitose, levando a crescimento do verme após alimentação e a crescimento do blastema, durante a regeneração.
Desde há muito se sabe que planárias podem resistir a jejum prolongado. Porém, Abeloos (1930) demonstrou que o jejum prolongado leva à diminuição de tamanho do verme, o que é acompanhado por regressão dos órgãos reprodutores. Assim, juntamente com a diminuição de tamanho, há diminuição da oviposição e da atividade locomotora. Surgem manchas irregulares do pigmento e o animal pode chegar à desintegração parcial ou total. Com retomada da alimentação normal, o indivíduo pode se recuperar, mas Abeloos lembrou que um animal reduzido por inanição não tem o mesmo poder de crescimento de um indivíduo jovem.
Bowen, Ryder e Dark (1976) com Polycelis tenuis objetivaram determinar os efeitos iniciais de privação na citoquímica e na organização funcional das células, nessa planária. Os autores constataram que há decréscimo do tamanho dos animais ao longo do tempo de privação e sugeriram que tal diminuição seria devida à ação continuada de vários fatores: redução das mitoses, aumento de morte celular, encolhimento celular, isso tudo levando à desdiferenciação celular. Mas, contrariamente a Abeloos, Bowen e seus colaboradores afirmaram que, aos 36 dias de privação, embora haja uma extensa perda de tecidos, todos os tipos de célula estão presentes na planária.
Davison (1973) observou que espécimes de Dugesia tigrina criados isoladamente uns dos outros em vasilhas com água e alimentação artificial diminuem de tamanho, ao passo que isso não ocorre com animais em grupo. Considerou que a diminuição talvez seja em função de esses animais isolados se dividirem em maior proporção do que os animais em grupo. Mas Davison apontou que talvez os animais isolados necessitem de condições de manutenção mais cuidadas do que os animais em grupo.
Além de observar o tamanho do animal, preocupamo-nos, também, em observar sua coloração.
Marcus (1964) descreveu que a cor de Dugesia tigrina varia quanto a tons e a intensidades, sendo que os diversos tipos de coloração e desenho são resultado da disposição do pigmento. Notou que animais provenientes de um mesmo local têm colorações e desenhos semelhantes. Marcus apontou que o grau de claridade do animal depende da alimentação. Nós, também, notamos que, após a alimentação com fígado cru, os animais ficam com uma coloração avermelhada e, pouco depois, mais escura do que a exibida antes da alimentação.
Método
As condições de manutenção dos animais
Neste estudo, observamos 27 planárias da espécie Dugesia tigrina, coletadas em tanques no campus da Cidade Universitária da Universidade de São Paulo, São Paulo, Capital, mantidas em três diferentes condições ambiente.
Nove planárias da condição de manutenção em grupo permaneceram, durante 35 dias consecutivos, juntas em um mesmo frasco com água, pedras e vegetação. Esporadicamente, a superfície da água do frasco era submetida a um jato de ar que a movimentava suavemente. O frasco era provido, duas vezes por semana, com nove caramujos do gênero Planorbis. Os animais eram, ainda, alimentados com pedaços de fígado cru de boi, oferecidos às planárias duas vezes por semana, em dias não coincidentes com aqueles nos quais os caramujos eram colocados no frasco.
Pedras, vegetação, caramujos, várias planárias e leve movimentação da água foram escolhidos como aspectos que fariam parte do ambiente de manutenção, por se considerar que são alguns dos elementos que usualmente compõem o habitat de Dugesia tigrina ou que têm semelhança com tais elementos.
Nove outras planárias foram mantidas durante os mesmos 35 dias isoladas umas das outras em nove frascos com água, compondo, assim, a condição de manutenção em isolamento.
Outras nove planárias permaneceram durante 35 dias na condição de manutenção mutável. Nessa condição de manutenção, as possibilidades de estimulação ambiente alteravam-se semana a semana. Assim, durante a primeira semana do estudo, as planárias da condição mutável permaneceram isoladas umas das outras em nove frascos com água. Na segunda semana, em cada recipiente, foi colocada uma pedra e vegetação. Na terceira semana, os animais, cada um em um recipiente com água, pedra e vegetação, passaram a receber, em duas vezes na semana, um caramujo do gênero Planorbis. Na quarta semana do estudo, os nove animais foram reunidos em um mesmo frasco com água, pedras e vegetação. Duas vezes por semana, eram colocados nove caramujos no frasco desses animais. Na quinta semana, à condição existente na semana anterior foi acrescentado um jato de ar que, esporadicamente, incidia na superfície da água, movimentando-a.
Os animais das condições em isolamento e mutável também recebiam alimento sob a forma de um pedaço cru de fígado de boi, colocado nos frascos duas vezes por semana.
Os recipientes nos quais ficaram as planárias durante o transcorrer do estudo eram de material plástico, incolor e transparente. Os frascos tinham formato de paralelepípedo, com as seguintes dimensões: 24 centímetros de comprimento, 12 centímetros de largura e sete centímetros de altura. Cada recipiente continha, aproximadamente, um litro de água mineral, sem gás.
Os frascos com as planárias foram mantidos em uma sala destinada apenas para fins do estudo, em ciclo controlado de luz-escuro, de 12 horas.
A situação de alimentação com fígado
Os pedaços de fígado oferecidos, duas vezes por semana, aos animais das três condições de manutenção eram colocados com auxílio de pinças sobre a base de cada recipiente, mais ou menos no centro da base. Procurávamos introduzir o fígado na água com o mínimo possível de perturbação e o fígado só era solto da pinça quando se achava apoiado na base do recipiente. O fígado permanecia nos frascos por um período de, aproximadamente, quatro horas. Em seguida, os frascos eram limpados. A limpeza consistia em retirar, de cada frasco, o pedaço de fígado, a(s) pedra(s) e a vegetação, caso houvesse, e toda a água. Em seguida, a superfície interna do frasco era limpada com pedaços de algodão envolvidos em palito e os elementos que haviam sido retirados eram recolocados nos frascos. Finalmente, punha-se água limpa em cada recipiente.
Na primeira semana do estudo, colocamos nos frascos pedaços de fígado de, aproximadamente, quinze gramas. Discutindo-se as observações realizadas, supusemos que as reações ao fígado, aparentemente pequenas em todos os grupos, talvez tivessem sido devidas ao tamanho reduzido dos pedaços de alimento.
Assim, na segunda semana, colocamos, para todos os animais, pedaços de fígado de 30 gramas, aproximadamente. Após nova discussão entre as observadoras, considerou-se que, embora a reatividade tivesse sido maior do que na semana anterior, ainda parecia pequena. Supôs-se, então, que tal fato estaria ocorrendo em função da forma como o fígado estava sendo colocado no frasco.
Desse modo, na terceira semana, embora mantido o mesmo tamanho do pedaço de fígado que na semana anterior, introduziu-se o fígado na água e, junto à base, a pinça com o fígado foi deslocada perpendicularmente duas vezes, com movimentos suaves. Pensamos que o movimento do fígado poderia contribuir para que o sumo se espalhasse mais rapidamente na água. Claro está que havia, no caso, também a presença de fatores associados ao movimento do fígado, como, por exemplo, projeção de sombras e movimento da água.
Visto terem-se verificado reações aparentemente claras, considerou-se, em discussão posterior à observação, que tal forma de apresentar o fígado deveria ser mantida nas duas semanas seguintes, de forma que tivéssemos um controle tal da situação que permitisse conclusões acerca das reações a esse tipo de alimentação.
As observações e os registros efetuados
Tendo em vista o objetivo deste estudo, cabe descrever o procedimento adotado para observação e registro de dados relativos a: (1) aspecto físico dos animais, compreendendo basicamente informações sobre comprimento, espessura, largura e coloração; (2) reprodução, implicando, aqui, em dados relativos a oviposição e número de filhotes; e (3) alimentação, considerando-se, especificamente, a situação de alimentação com fígado.
Para fazer parte do grupo de 27 animais estudados foram escolhidas, dentre as planárias coletadas, aquelas que tinham comprimentos em torno de 14 milímetros, que estavam intactas e que tinham padrões de coloração semelhantes entre si.
Para medir o comprimento das planárias, procedíamos da seguinte forma: colocávamos os animais individualmente em placas de Petri, com água, apoiadas sobre um pedaço de papel milimetrado. Quando a planária estivesse exibindo locomoção por deslizamento com manutenção da postura em seta (Dolci, 1978) procedíamos à mensurarão, contando os quadrados ocupados pelo animal, no sentido de seu comprimento.
Além de os animais terem sido medidos imediatamente antes do início do estudo, foram-no, também, após o seu término.
Diariamente, procedia-se à contagem dos animais e registravam-se mortes porventura ocorridas. Se necessário, eram também anotados dados qualitativos acerca da aparência física das planárias.
No que se refere à reprodução, dia a dia, eram registrados o número total de cápsulas depositadas, bem como, o número de filhotes, caso houvesse.
Na situação de alimentação com fígado os animais eram observados, uma vez por semana, antes e depois do fígado ser introduzido nos frascos. A observação tinha duração de trinta minutos e, concomitantemente, efetuavam-se registros de comportamento. Transcorridos 15 minutos desde o início da observação, a mesma era interrompida e colocavam-se os pedaços de fígado cru de boi nos recipientes, realizando-se, a seguir, os outros 15 minutos de observação.
Durante as observações registrava-se por amostragem de tempo, a intervalos de um minuto, o número de animais que exibiam cada uma de três categorias de comportamento que diziam respeito a atividade motora dos animais: estacionamento, locomoção e deslocamentos de parte anterior. Quando fosse o caso, eram registrados esses mesmos comportamentos, porém, em relação ao fígado: estacionamento sobre o fígado; locomoção sobre o fígado e deslocamento de parte anterior sobre o fígado. Descrições detalhadas dessas categorias de comportamento encontram-se em outro texto (Dolci-Palma, 1985).
Todos os registros foram efetuados por pessoas com prática em observação de comportamento de planárias e no tipo de registro aqui utilizado.
Resultados e discussão
Consideraremos, inicialmente, em termos dos resultados obtidos, aqueles referentes ao aspecto físico dos animais observados.
A Tabela I mostra os dados relativos ao comprimento em milímetros das planárias de cada uma das três condições de manutenção, no início e no final do estudo, ou seja, no trigésimo quinto dia de observação. Lembremos que, apenas na condição em isolamento, havia possibilidade de reconhecermos individualmente os animais desde o primeiro e até o último dia da investigação; assim, na tabela, os dados relativos a um mesmo animal da condição em isolamento, nos dois momentos do estudo, encontram-se alinhados.
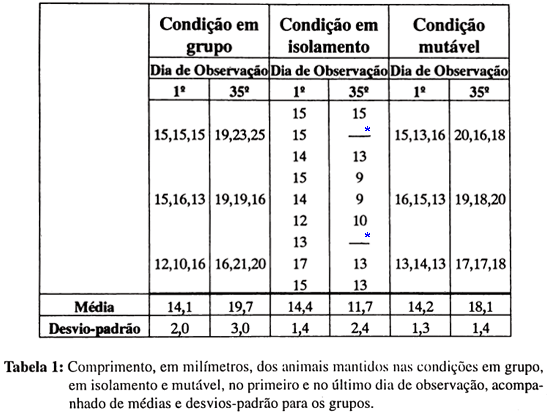
Notamos, na Tabela I, que houve aumento em comprimento das planárias da condição em grupo, se for considerado o conjunto de animais. Como dado qualitativo, cabe apontar que essas planárias, no final do estudo, mostraram-se mais largas, mais espessas e mais escuras do que no início.
No grupo de planárias mantidas na condição em isolamento ocorreram duas mortes, uma no décimo segundo dia do estudo e outra no trigésimo segundo dia. Observamos, na Tabela I, que apenas um animal conservou o mesmo comprimento do início até o final das observações; todos os demais diminuíram de comprimento, perdendo de um a seis milímetros. No final do estudo, constatamos que todos os animais desse grupo estavam mais claros, menos espessos e mais estreitos do que no início.
O grupo de animais mantido na condição mutável teve seu comprimento aumentado do início para o final do estudo, como podemos constatar na Tabela I. Pudemos também notar que os animais se tornaram, ao longo do estudo, mais largos, mais espessos e mais escuros.
Assim, pudemos concluir que o ocorrido com a aparência física dos animais da condição mutável foi bastante semelhante ao ocorrido com aqueles da condição em grupo e diverso do acontecido com as planárias da condição em isolamento.
Valendo-nos da comparação entre as condições de manutenção em isolamento e em grupo, podemos supor, em um primeiro momento, que o aumento em proporções desses últimos animais estaria relacionado à possibilidade de ingerir caramujos, uma vez que os animais em isolamento não tiveram caramujos à disposição, para caça, ao longo de todo o estudo, contrariamente às planárias em grupo. Lembremos os estudos de Baguna (1974, 1976) mostrando que o aumento em proporções físicas de planárias de água doce é função da quantidade e da qualidade da alimentação. Quando comparada a condição mutável às outras duas condições de manutenção, pode-se sugerir, de modo ainda mais enfático, que a possibilidade de caçar e ingerir caramujos - que passou a ser oferecida às planárias da condição mutável a partir da terceira semana do estudo - tivesse sido um fator a contribuir em muito para o aumento em proporções dos animais da condição em grupo e da condição mutável. E, de fato, ao longo do estudo, pudemos notar que as planárias, tanto na condição de manutenção em grupo quanto na condição mutável, caçavam e ingeriam caramujos quando estes eram colocados à disposição nos frascos. A análise detalhada de como as planárias se comportavam em relação aos caramujos acha-se publicada em outro local (Dolci-Palma, 1987).
Em um segundo momento, e considerando as conclusões de Baguna, antes referidas, há outro aspecto a ser considerado. Ao longo do estudo, observamos diferenças e semelhanças entre os grupos de planárias na forma como reagiram aos pedaços de fígado colocados nos frascos.
Entre os animais em grupo, quando o fígado era oferecido às planárias, a maioria deixava de exibir-se em estacionamento, apresentava alguma atividade motora geral - na forma de locomoção e de deslocamentos da parte anterior - e, também, dirigida especificamente ao fígado - na forma de locomoção sobre o fígado e de deslocamentos da parte anterior sobre o fígado e ao redor dele - e logo passava a comer, ou seja, a exibir-se em estacionamento sobre o fígado (ver Figura 1). O primeiro animal a entrar em contato com o alimento o fazia logo nos primeiros minutos após a introdução do fígado na água. Tal contato, depois de iniciado, era sempre ininterrupto e, no último minuto da observação, pelo menos sete dos nove animais do grupo haviam entrado em contato com o fígado. Essa forma de reação mostrou-se muito semelhante nas três ocasiões em que o fígado era ligeiramente balançado quando submerso na água do frasco, ou seja, nas três semanas finais do estudo.
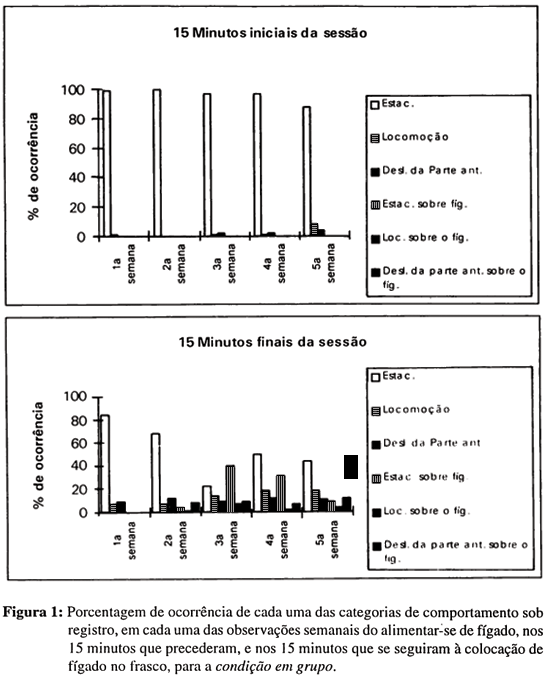
Na primeira semana, quando o pedaço de fígado era menor e apenas depositado no frasco, sem ser balançado, nenhum animal entrou em contato com o alimento, durante os 15 minutos finais da observação (ver Figura 1). Na segunda semana, quando o pedaço de fígado era maior, embora colocado no frasco sem ser balançado, as reações ao fígado foram pequenas (ver Figura 1).
Assim, o ter-se aumentado em tamanho o pedaço de alimento posto no frasco parece ter tido um pequeno efeito sobre a reatividade ao alimento. Já o aumento em tamanho, quando associado ao movimentar-se do fígado na água, foi um fator relevante para o desencadeamento das reações ao alimento, nesse grupo de animais.
Na Figura 2, temos resultados relativos à alimentação com fígado para o grupo de planárias da condição em isolamento. A incidência de quaisquer categorias registradas, à exceção de estacionamento, foi baixa: na maior parte das vezes com porcentagens menores do que 15 por cento. Contudo, quando comparadas entre si as cinco semanas do estudo, vemos que a reatividade ao fígado foi ligeiramente maior nas três últimas semanas do estudo do que nas duas primeiras, sugerindo um possível efeito do movimento de balançar o fígado na água sobre a ativação dos animais e a reatividade ao alimento.
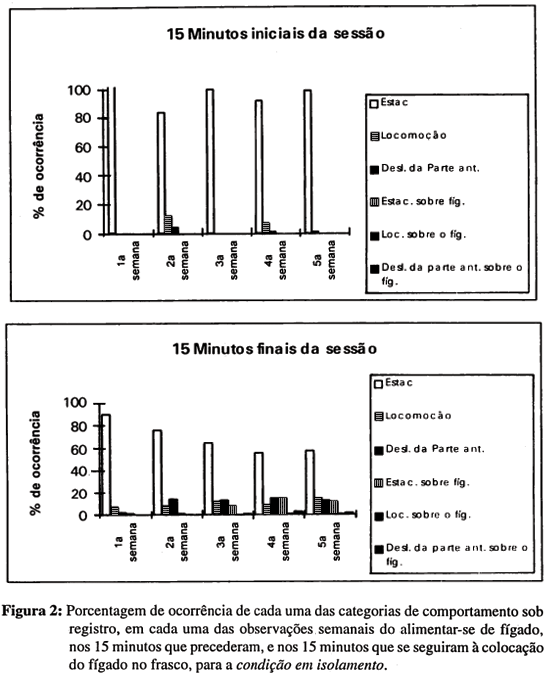
Assim, os animais da condição em isolamento podiam deixar de exibir estacionamento quando o fígado era colocado com balanço nos frascos, mas nem todos aqueles que o faziam eram vistos em contato com o fígado. Menos da metade dos animais chegava a entrar em contato com o alimento e foi visto interromper-se o contato depois que o animal já se encontrava estacionado sobre o alimento. Quando deixavam de estar estacionados, os animais exibiam locomoção e deslocamentos da parte anterior, mas podiam depois voltar a estacionar sem entrar em contato com o alimento.
Na Figura 3 vemos que, nas duas primeiras semanas da investigação, quando o fígado não era balançado no momento de sua introdução nos frascos dos animais da condição mutável, praticamente não se alteraram as categorias de comportamento da primeira para a segunda metade da observação, à exceção de deslocamentos da parte anterior, na segunda semana. Vemos também que, quando o fígado passou a ser introduzido com movimento nos frascos, os comportamentos dos animais diferiram visivelmente daqueles exibidos quando o fígado era introduzido sem movimentação.
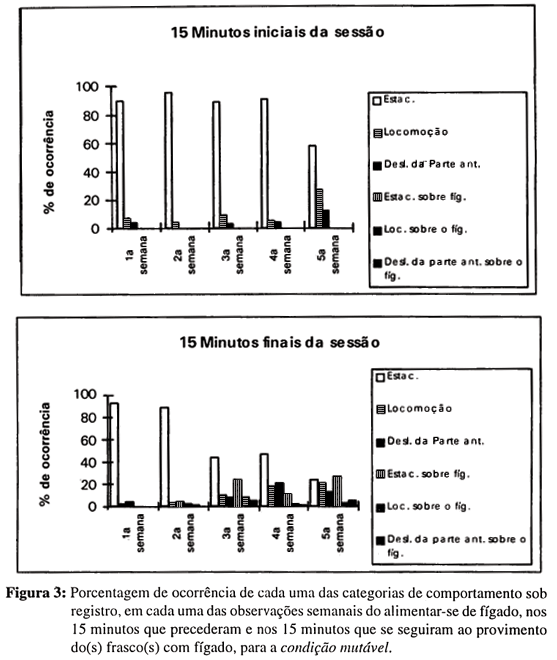
Nessas três oportunidades, os animais da condição mutável deixaram de exibir estacionamento, se não se encontrassem já em mobilidade (ver Figura 3). De três a oito planárias entravam em contato ininterrupto com o alimento após, no máximo, três minutos depois do provisionamento dos frascos com o fígado. Em geral, depois de saírem de estacionamento, as planárias aumentavam a atividade motora geral e nenhum animal, após ter entrado em contato com o fígado, voltou a estacionar fora dele.
Portanto, os dados relativos à reatividade ao fígado foram semelhantes entre as planárias em grupo e aquelas da condição mutável e ambos os conjuntos de resultados diferiram do modo de reação apresentado pelas planárias em isolamento.
No que se refere à reprodução, nenhum dos animais observados exibiu fissão transversal, o que está de acordo com as observações de Molina (1976) que, estudando Dugesia tigrina coletada nos mesmos locais onde foram colhidas as planárias aqui estudadas, não observou a divisão transversal como processo reprodutivo.
Quanto à deposição de cápsulas ao longo do estudo, a Tabela II mostra, para cada uma das três condições de manutenção estudadas, a taxa diária de postura de cápsulas para cada uma das semanas do estudo. Os animais da condição em grupo mostraram certa regularidade na oviposição ao longo dos 35 dias. No final do trabalho, haviam sido postas em torno de 69 cápsulas e haviam sido retiradas do frasco um total de 120 planarinhas, que começaram a ser vistas no recipiente a partir da quarta semana do estudo.
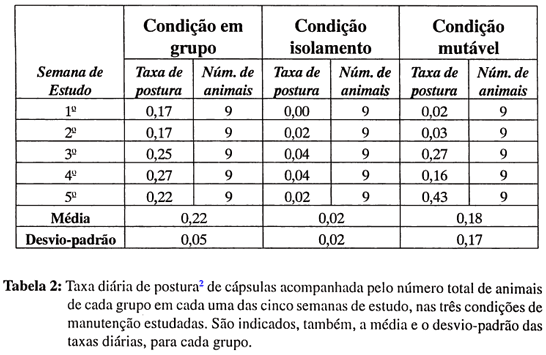
Apenas um animal da condição em isolamento depositou cápsulas ao longo da investigação e o fêz num total de seis cápsulas, sendo que nenhuma delas eclodiu até o final das observações.
Entre os animais da condição mutável, não foi observada eclosão de cápsulas e houve deposição das mesmas ao longo de todo o estudo, porém, com menor regularidade, semana a semana, do que entre os animais mantidos na condição em grupo.
Assim, na condição mutável, na primeira e na segunda semanas do estudo, a taxa de postura foi semelhante àquela calculada para o grupo em isolamento (ver Tabela II). Lembremos, que, nessas duas semanas, a condição mutável era bastante semelhante àquela em isolamento no que se referia a condições do ambiente, o que sugere que pedras e vegetação não têm um efeito desencadeador na oviposição. Na terceira semana, quando caramujos eram colocados à disposição para caça e ingestão, a taxa de postura de cápsulas, para as planárias da condição mutável, tornou-se semelhante àquela da condição em grupo, diferindo daquela da condição em isolamento. Esse dado sugere uma relação de dependência entre oviposição e possibilidade de caçar e ingerir presas vivas. Na quarta semana do estudo, quando os animais da condição mutável passaram a estar reunidos em um mesmo frasco, com caramujos à disposição, diminuiu a taxa de postura em relação à condição em grupo e, na quinta semana, aumentou. A taxa diminuída na quarta semana poderia ser a expresso de uma relativa inibição de postura provocada pela reunião dos animais em um mesmo frasco e/ou o resultado de ocorrerem atividades incompatíveis com a postura. Já na quinta semana, a taxa foi maior, talvez como um efeito compensatório da redução havida na quarta semana.
Na tentativa de compreendermos as semelhanças e diferenças entre os grupos de planárias estudadas no que diz respeito à reatividade ao fígado, à reprodução e ao aspecto físico dos animais, consideraremos as implicações contidas em cada uma das três formas de manter as planárias.
Podemos dizer que as planárias da condição de manutenção em grupo estavam em uma situação privilegiada: o alimento (sob a forma do pedaço de fígado colocado no recipiente, ou de caramujos) era abundante; os animais estavam protegidos de qualquer condição de predação; a presença de pedras e vegetação constituía uma proteção à iluminação ambiente e possibilitava estimulação mecânica e química diversificada; e, ainda, a situação era estável no que respeitava às condições do ambiente, semana a semana. Nessa medida, a situação da condição em grupo pode ser considerada como superprotegida e artificial. Porém, dentro dos limites de duração do estudo, podemos concluir também que se tratou de uma situação bastante semelhante àquela presente no habitat de Dugesia tigrina e conveniente à manutenção dessa planária em laboratório, propiciando condições favoráveis à sobrevivência dos indivíduos e à preservação do grupo.
No caso da condição de manutenção em isolamento, temos uma situação desfavorável à sobrevivência dos indivíduos, que os levou a um estado de depauperamento físico e impossibilitou-os de exibirem certos comportamentos que são típicos de planárias. Considerada especificamente a reatividade ao fígado, houve mesmo modificação no caráter de alguns comportamentos, relativamente à forma como foram mostrados pelos animais das condições em grupo e mutável.
Lembremos que a condição em isolamento era mais estável e padronizada do que a condição em grupo e que os animais estavam em uma situação relativamente mais pobre de estimulação, não tendo, sequer, uma forma de proteção eficaz em relação à estimulação luminosa ambiente.
Na condição mutável, os dados relativos à reatividade ao fígado e ao aspecto físico dos animais mostraram uma tendência semelhante aos dados correspondentes no caso da condição em grupo. Nessa medida, a menor estabilidade das condições existentes para esse terceiro grupo de planárias parece não ter sido um fator relevante em diferenciá-los dos animais na condição em grupo. Quanto à oviposição, as diferenças observadas em relação aos animais em grupo foram hipoteticamente relacionadas à menor estabilidade da condição de manutenção mutável.
Em conclusão, os resultados obtidos quanto ao aspecto físico dos animais, à reprodução e à reatividade ao fígado indicam que as condições de manutenção geram um conjunto de efeitos diferenciados nas planárias assim mantidas por um determinado espaço de tempo.
Estudos experimentais deveriam ser feitos com o objetivo de concluir acerca da contribuição relativa que cada um dos fatores presentes ou ausentes em cada uma das condições de manutenção aqui empregadas teria tido para o modo característico como se comportaram e se apresentaram os animais estudados. Assim, teríamos condições de estabelecer, com maior segurança, qual dentre os seguintes fatores seria o responsável por tal modo de se comportar: Isolamento social? Depauperamento físico e/ou fisiológico? Problemas com o alimentar-se? Problemas com locais adequados para oviposição e abrigo? Estabilidade maior ou menor da situação de manutenção? Pobreza relativa de estimulação ambiente? Ou seriam responsáveis alguns ou todos os fatores combinados?
REFERÊNCIAS BIBLIOGRÁFICAS
ABELOOS, M. Recherches expérimentales sur la croissance et la régéneration chez les Planaires. Bulletin Biologique de la France et de la Belgique, v.64, p.1-140, 1930. [ Links ]
BAGUNA, J. Dramatic mitotic increase in planarians after feeding, and a hypothesis for the control mechanism. Journal of Experimental Zoology, v.190, p.117-22, 1974. [ Links ]
BAGUNA, J. Mitosis in the intact and regenerating planarian Dugesia mediterranea n. sp.: mitotic studies during growth, feeding and starvation. Journal of Experimental Zoology, v.195, n.1, p.53-64, 1976. [ Links ]
BELLAMY, L.S.; REYNOLDSON, T.B. Behaviour in competition for food amongst lake-dwelling Triclads. Oikos, v.25, p.356-64, 1974. [ Links ]
BOWEN, I.D.; RYDER, T.A.; DARK, C. The effects of starvation on the planarian worm Polycelis tenuis Iijima. Cell and Tissue Research, v.169, n.2, p.193-210, 1976. [ Links ]
CARNEY, R.E.; MITCHELL, G.L. Reactions of planarians after cannibalization of planarians exposed to four stimulus combinations. Journal of Biological Psychology, v.20, n.2, p.44-9, 1978. [ Links ]
CORNING, W.C. Retention of a position discrimination after regeneration in planarians. Psychonomic Science, v.5, n.1, p.17-8, 1966. [ Links ]
DAVISON, J. Population growth in planaria Dugesia tigrina (Girard): Regulation by the absolute number in the population. Journal of General Physiology, v.61, n.6, p.767-85, 1973. [ Links ]
DOLCI, I.A. Um estudo sobre o comportamento de planárias em situação padronizada. São Paulo, 1978. 204p. Dissertação (Mestrado) - Instituto de Psicologia, Universidade de São Paulo. [ Links ]
DOLCI-PALMA, I.A. Caça e ingestão de presas em planárias da espécie Dugesia tigrina mantidas em laboratório. Ciência e Cultura, v.39, n.5/6, p.557-60, 1987. [ Links ]
DOLCI-PALMA, I.A. A Planária no ambiente de manutenção: um estudo biológico e psicológico. São Paulo, 1985. 2v. Tese (Doutorado) - Instituto de Psicologia, Universidade de São Paulo. [ Links ]
GALTSOFF, P.S.; LUTZ, F.E.; WELCH, P.S.; NEEDHAM, J.C. Culture methods for invertebrate animals. New York, Dover, 1959. [ Links ]
JACOBSON, A.L.; HOROWITZ, S.D.; FRIED, C. Classical conditioning, pseudoconditioning or sensitization in the planarian. Journal of Comparative and Physiological Psycology, v.64, n.1, p.73-9, 1967. [ Links ]
LEVISON, M.J.; GAVURIN, E.I. Truly randon control in Pavlovian conditioning of planaria (D. dorotocephala). Psychological Reports, v.45, n.3, p.987-92, 1979. [ Links ]
LOOMIS, L.L.; NAPOLI, A.M. Transfer of training through two cannibalisms of planarian. Journal of Biological Psychology, v.17, n.1, p.37-40, 1975. [ Links ]
McCONNELL, J.V.; JACOBSON, A.L.; KIMBLE, D.P. The effects of regeneration upon retention of a conditioned response in the planarian. Journal of Comparative and Physiological Psychology, v.52, p.1-5, 1959. [ Links ]
MARCUS, E. Sobre Turbellaria límnicos brasileiros. Boletim da Faculdade de Filosofia, Ciências e Letras, Universidade de São Paulo: Zoologia, v.11, p.5-254, 1964. [ Links ]
MASON, P.R. Chemo-klino-kinesis in planarian food location. Animal Behaviour, v.23, p.460-9, 1975. [ Links ]
MICKLIN, M.P.; MAY, J.C. An apparent frustration effect in planarians. Journal of Biological Psychology, v.17, n.2, p.4-9, 1975. [ Links ]
MOLINA, F.M.L.R. Reprodução e ciclo biológico de Dugesia tigrina (Girard, 1850) (Turbellaria, Tricladida, Paludicola). São Paulo, 1976. 103p. Dissertação (Mestrado) - Instituto de Biociências, Universidade de São Paulo. [ Links ]
PEARL, R. The movements and reactions of fresh-water planarians: a study in animal behaviour. Quarterly Journal of Microscopical Science, v.46, p.509-714, 1903. [ Links ]
REYNIERSE, J.H.; GLEASON, K.K.; OTTEMAN, R. Mechanisms producing aggregations in planaria. Animal Behaviour, v.17, n.1, p.47-63, 1969. [ Links ]
SILVA, P.K. Evidence for aggregation from a field study of a flatworm population. Archiv fuer Hydrobiologie, v.81, n.4, p.493-507, 1978. [ Links ]
SILVA, P.K. The factors affecting the feeding of Dendrocoelum lacteum (Müller) (Turbellaria, Tricladida) on Asellus acquaticus. Archiv fuer Hydrobiologie, v.77, p.347-74, 1976. [ Links ]
SIMPSON, G.G. The study of evolution: methods and present status of theory. In: ROE, A.; SIMPSON, G.G., eds. Behavior and evolution. London, Yale University Press, 1958. p.7-26. [ Links ]
STEWART, A.M. Phototaxis in three freshwater triclad flatworms. American Zoologist, v.8, p.745, 1968. /Abstract/ [ Links ]
WALTER, H.E. The reaction of planarians to ligth. Journal of Experimental Zoology, v.5, n.1, p.35-162, 1907. [ Links ]
WULZEN, R.; BAHRS, A.M. Unbalance in planarian nutrition. Physiological Zoology, v.4, p.204-13, 1931. [ Links ]
YAREMKO, R.M.; KIMMEL, H.D. Two procedures for studying partial reinforcement effects in classical conditioning of the planarian. Animal Behaviour, v.17, n.1, p.40-2, 1969. [ Links ]
* Morte do animal.
1 Parte da tese de Doutorado em Psicologia - área de concentração: Psicologia Experimental - IPUSP, 1985. Orientador: Walter Hugo de Andrade Cunha.
2 A taxa foi calculada da mesma forma que em Molina30: (número de cápsulas por semana / número de dias na semana em que foram postas cápsulas), número de vermes.
