










Serviços Personalizados
artigo
 Português (pdf)
Português (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
Indicadores

Compartilhar

 Permalink
PermalinkPsicologia USP
versão On-line ISSN 1678-5177
Psicol. USP v.17 n.4 São Paulo dez. 2006
ARTIGOS ORIGINAIS
Espaço de cores1
Color space
Espace de couleur
Claudia Feitosa-SantanaI,2; Nestor Norio OiwaI,3; Marcelo Fernandes da CostaI,4; Klaus Bruno TiedemannI,4; Luiz Carlos de Lima SilveiraII,5; Dora Fix VenturaI,4
I Universidade São Paulo - USP
II Universidade Federal do Pará - UFPA
RESUMO
O artigo apresenta definições para os termos espaço de cores e sistemas de cores; classifica, de acordo com David Brainard (2003), os sistemas de cores em dois grupos: aparência de cores e diferenças de cores. Dentre os diversos sistemas de cores existentes, o artigo descreve dois deles: o sistema de cores Munsell &– um dos mais utilizados entre os sistemas de aparência de cores &– e a descrição do sistema de cores CIE 1931 &– um dos mais utilizados dentre os sistemas de diferença de cores. Faz-se uma retrospectiva histórica da busca por espaços de cores que representem a percepção de cores humana assim como as diversas reconstruções de espaços de cores por métodos eletrofisiológicos ou psicofísicos. Muitas dessas reconstruções utilizam a escala multidimensional (mds). O artigo também introduz a possibilidade da reconstrução dos espaços de cores de pacientes com discromatopsia adquirida como uma distorção do espaço de indivíduos tricromatas normais.
Palavras-chave: Percepção de cor. Discriminação de cores. Psicofísica. Escala multidimensional.
ABSTRACT
The article presents definitions for the terms color space and color system. In agreement with David Brainard (2003), it classifies the color systems in two groups: color appearance and color difference. Amongst the existing color systems, the article also presents the description of the Munsell Color System, one of the most used among the color appearance systems, and of the CIE 1931, one of the most used color difference systems. In addition, an historical retrospect of the search for color spaces that represent the human color perception as well as the reconstruction of color spaces with electrophysiological and psychophysical methods, are described. Many of these reconstructions use the multidimensional scaling methods (mds). Finally, the article presents the possibility for reconstruction of color spaces of patients with acquired dyschromatopsia as a distortion of the color space of normal trichromats.
Keywords: Color perception. Color discrimination. Psychophysics. Multidimensional scaling.
RÉSUMÉ
L'article présente des définitions de l'espace de couleur et du système de couleur; selon David Brainard (2003), il classifie les systèmes de couleur en deux groupes: l'aspect de couleur et la différence de couleur. Parmi les systèmes de couleur existants, l'article présente la description de deux d'entre eux: le Système de Couleur de Munsell, l'un des plus utilisés parmi les systèmes d'aspect de couleur, et la description de CIE 1931, l'un des systèmes de différence de couleur les plus utilisés. L'article présente une rétrospective historique de la recherche des espaces de couleur qui représentent la perception humaine de la couleur aussi bien que diverses reconstructions des espaces de couleur, par l'emploi des méthodes d'électrophysiologie et de psychophysique. Plusieurs de ces reconstructions emploient l'échelle multidimensionnelle (mds). L'article présente la possibilité de la reconstruction des espaces de couleur chez les patients avec dyschromatopsie acquise comme une déformation des espaces de couleur trichromatiques normaux.
Mots-clés: Perception de couleur. Discrimination de couleur. Psychophysique. Échelle multidimensionnelle.
Definindo a percepção de cores
A luz, ao atravessar os meios ópticos oculares, excita moléculas fotossensíveis dos fotorreceptores da retina que, por sua vez, iniciam o processo de codificação da informação presente nos raios luminosos até que após extenso processamento neural em vários níveis da retina, do tálamo e do córtex cerebral, ocorre a percepção da cor (Backhaus, 1998).
Para Hering (1874), a percepção visual é formada a partir de seis cores elementares: verde, vermelho, azul, amarelo, branco e preto. A percepção do branco é o resultado da combinação particular de cores elementares. O preto é, de fato, uma percepção de cor, uma vez que a ausência de percepção não tem significado neural ou psicológico.
Definindo o termo espaço de cores
Há diversas definições para espaço de cores na literatura científica (Backhaus, 1998; Brainard, 2003; Helm, 1964; Indow, 1995; Paramei, 1996; Shepard & Carroll, 1966). Neste texto, utilizou-se a expressão espaço de cores em três situações: na representação gráfica das relações entre as cores, o que inclui os desenhos esquemáticos da Antigüidade; nos espaços de cores obtidos por experimentos fisiológicos; e nos espaços de cores perceptuais construídos com os resultados de métodos psicofísicos.
Definindo o termo sistema de cores
Existem diversos métodos de sistematização do espaço de cores e, de acordo com Brainard (2003), essa sistematização pode ser feita para a especificação da aparência das cores ou da diferença entre as cores, mas ainda não há um sistema de cores perfeito para nenhuma das duas finalidades. Dentre os sistemas de aparência de cores, o mais conhecido e utilizado é o Sistema de Cores de Munsell (Munsell Color System). Grande parte dos sistemas de diferenças de cores foi desenvolvida a partir daquele criado pela Comission Internationale de l'Eclairage de 1931 (CIE 1931) (Brainard, 2003).
A retrospectiva histórica dos espaços e sistemas de cores
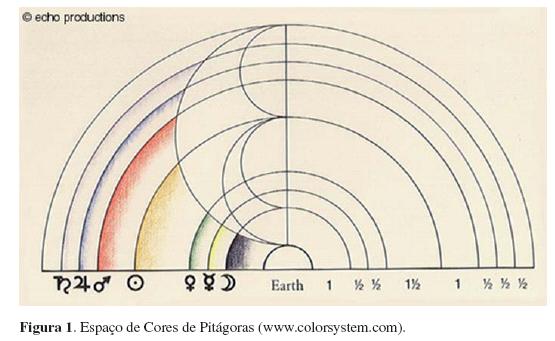
Entre os primeiros registros de espaços de cores, pode-se destacar o trabalho de Pitágoras (circa 570-500 a.C.), que criou um espaço de cores semicircular relacionando as notas da escala musical de tons e meio tons aos planetas que, por sua vez, eram representados por determinadas cores (Figura 1) (Fisher, 1999).
Aristóteles (384-322 a.C.) apresentou seu espaço de cores em uma seqüência linear, do branco ao preto, compreendendo cinco cores entre elas. Nesse estudo, a luz branca da Lua tinge-se de amarelo, ao longo do dia torna-se laranja e, em seguida, vermelho, violeta, verde e azul, chegando, então, ao preto, que representa a escuridão da noite. Aristóteles foi, provavelmente, o primeiro a investigar a mistura de luzes quando observou a formação do verde em uma parede de mármore branco, após a passagem de um feixe de luz branco por dois fragmentos de vidro nas cores azul e amarelo, respectivamente (Fisher, 1999).
O inglês Robert Grosseteste (circa 1170-1253 d.C.), primeiro Chancellor da Universidade de Oxford, apresentou seu espaço de cores no livro De Colore, de 1230. Neste espaço, o branco (“albedo” ou “lux clara”) e o preto (“nigredo” ou “lux obscura”) estão separados das demais cores, localizados em dois extremos do mesmo eixo, como a maioria dos espaços de cores utilizados atualmente (Fisher, 1999). De acordo com Gage (2001), a seqüência do branco ao preto, que determina uma escala de cinza, foi anteriormente demonstrada por Avicenna (980-1037).
Por volta de 1435, Leon Battista Alberti (1404-1472), em seu livro Della Pintura, apresenta um espaço de cores baseado em duas oponências cromáticas: verde-vermelho e azul-amarelo. Essas duas oponências cromáticas constituíram, séculos mais tarde, o cerne da teoria de oponência de cores de Ewald Hering (1839) e tiveram suas bases funcionais comprovadas por métodos eletrofisiológicos por (De Valois, Abramov, & Mead, 1967). Alberti inseriu, também, a forma tridimensional da representação dos espaços de cores, embora ainda não fosse baseada em arranjos circulares (Fisher, 1999; Mollon, 2003).
No início do século XVII (1611), o astrônomo finlandês Aron Sigfrid Forsius (também conhecido como Siegfried Aronsen; circa 1550-1637), um dos precursores desse campo, apresentou um espaço de cores em forma de círculo e, a partir do mesmo, estabeleceu um espaço esférico (Fisher, 1999; Gage, 2001; Parkhurst & Feller, 1982).
Pouco depois, em 1613, o jesuíta belga Franciscus Aguilonius (1567-1617) incluiu em sua teoria um espaço de cores que foi, provavelmente, o primeiro deles concebido com as três cores-pigmentos primárias: “flavus, rubeus e caeruleus” (amarelo, vermelho e azul). As ilustrações de seu trabalho foram feitas por Paul Rubens que recebeu forte influência desta teoria de cores em suas obras (Campenhausen, 2001; Fisher, 1999; Jaeger, 1984).
O médico inglês Francis Glisson (1597-1677) apresentou, em 1677, um trabalho sólido baseado nas primárias vermelho, amarelo e azul da mistura de cores, em uma escala de cinzas composta por 23 etapas entre o branco e o preto. Juntamente com Forsius, Glisson também é considerado um precursor dos sistemas de cores (Fisher, 1999; Gage, 2001; Kuehni & Stanziola, 2002).
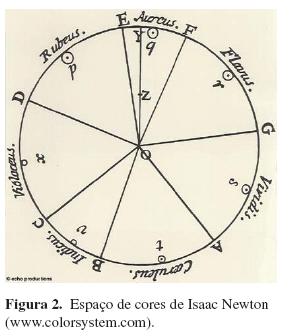
Isaac Newton (1642-1727), com o auxílio de um prisma, comprovou no Experimentum Crucis, que as cores-luzes, nomeadas por ele como “rubeus, aurantius, flavus, viridis, caeruleus, indicus e violaceus” (vermelho, alaranjado, amarelo, verde, azul, índigo e violeta), são cores componentes da luz branca. Em 1704, Newton publicou seu primeiro livro, Opticks, apresentando o espaço de cores em forma de círculo (Figura 2). Trata-se do disco de Newton: a rotação de um disco iluminado com luz branca e pintado com a seqüência de cores do espaço de Newton resulta, para o observador, em um disco branco. A partir desse momento, todos os outros espaços de cores não mais incluiriam o branco e o preto dentre as cores do espectro visível. Em seu espaço de cores, as cores localizadas na borda da circunferência, as quais correspondem às cores do espectro luminoso, sendo essa região então chamada de locus espectral, estão representadas em sua maior saturação ou pureza do matiz. Conforme se aproximam do centro da circunferência conservam o matiz, mas vão incorporando quantidades crescentes de branco, reduzindo a saturação do matiz. Apesar da grande maturidade na elaboração deste espaço, Newton incorporou uma analogia com a escala musical e não se preocupou com uma maior elaboração sistemática das cores. Em seu espaço não puderam ser representadas as gamas de púrpuras que são compostas pela mistura das cores das duas pontas da refração do prisma, mas Newton deixou por escrito uma referência a essa limitação (Mollon, 2003; Paramei, 2004).
Ao longo do século XVIII, principalmente na sua segunda metade, muitos investigadores deram contribuições importantes para esse campo de conhecimento o que permitiu que Thomas Young pudesse sintetizar a teoria tricromática do sistema visual humano e, assim, refletir nas elaborações dos espaços de cores que teriam por finalidade reproduzir a percepção cromática humana. No ano de 1755, Charles Bonnet (1720-1793) mencionou a existência de ressoadores de luz na retina. Curiosamente, em 1760, Bonnet descreveu um quadro de alucinações de seu pai, que possuía, aos 87 anos, problemas visuais em decorrência, provavelmente, de catarata e que hoje é conhecida como Síndrome Charles Bonnet. Em 1756, Mikhail Vasilevich Lomonosov sugeriu, como complemento da tricromacia física da luz, a tricromacia fisiológica: três tipos de partículas presentes na membrana escura do fundo do olho. Em 1775, anos depois da morte do matemático e astrônomo alemão Tobias Mayer (1723-1762), Georg Christoph Lichtenberg publicou uma aula na qual Mayer apresentou as três cores-luzes primárias que o olho humano seria capaz de perceber: vermelho, amarelo e azul (Mollon, 2003).
No último quarto do século XVIII ocorreram as duas últimas contribuições fundamentais. George Palmer (1740-1795), um especialista na fabricação de vidros coloridos, no ano de 1777, propôs a existência de três tipos de luzes que corresponderiam, respectivamente, a três tipos de partículas, às vezes nomeadas por ele como moléculas ou membranas, na retina. Também em Londres, a menos de um quilômetro de distância, John Elliot (1747-1787), em 1780, aventou que os órgãos sensoriais humanos, através de ressoadores, poderiam ser ativados mecanicamente na presença de estímulos apropriados. Elliot também teve a capacidade de intuir que o sistema visual humano não possuiria ressoadores para todas as freqüências existentes (Mollon, 2003).
Dessa forma, todos os conceitos fundamentais para a elaboração da teoria tricromática já estavam disponíveis no final do Século XVIII, mas sua elaboração só foi alcançada em 1801-02 &– mesmo ainda não tendo adquirido o nome de tricromacia &– por Thomas Young (1773-1829), em uma palestra na Royal Society de Londres. Com os conhecimentos adquiridos em suas pesquisas no campo da acústica, sua teoria sobre as cores foi ganhando firmeza e se apresentou de forma sólida, em 1817, em um artigo de sua autoria: Chromatics, elaborado para a Encyclopaedia Britannica. Young concluiu que diferentes comprimentos de onda correspondiam a diferentes matizes; os mais curtos se apresentavam em violeta e os mais longos em vermelho. Demonstrou, ainda, com bastante precisão, um mapa de distribuição dos matizes do espectro visível e, quando convertemos as frações de polegadas para nanômetros, vemos que suas estimativas eram próximas das reais dimensões que hoje conhecemos (Mollon, 2003).
De acordo com Mollon (2003), alguns autores atribuem a Young a compreensão e descrição do fenômeno da constância de cores, comumente utilizada na língua inglesa como color constancy. Young realmente descreveu esse fenômeno, além de interligá-lo ao contraste simultâneo das cores, mas o primeiro relato de que se tem conhecimento data de 1694, tendo sido feito por Philippe De La Hire (1640-1718), que afirmou o fato de não percebermos que as cores são diferentes sob a luz do dia ou sob a luz de velas. Ainda de acordo com Mollon (2003), o artigo mais brilhante a respeito da constância de cores foi feito em 1789, poucas semanas antes da revolução francesa, na Academia Real de Ciências de Paris, por Gaspard Monge (1746-1818). Em sua conferência, vestindo uma malha vermelha, Monge pediu a seus colegas que a observassem através de uma lente vermelha. Surpresos, os presentes tiveram a sensação de que a malha tinha um vermelho esbranquiçado, praticamente branco6. Monge tinha consciência de que essa sensação era mais forte nas cenas com brilho mais intenso e relacionou tal observação a um segundo fenômeno, o das Sombras Coloridas:7
Se um ambiente for iluminado pela luz do sol que passa através de uma cortina vermelha que tem um buraco, o feixe de luz que passar pelo buraco e cair sobre um pedaço de papel não irá parecer vermelho e sim “bastante verde”.
Pouco tempo depois das contribuições de Young e suas observações a respeito do contraste simultâneo de cores, o químico francês Michel Eugène Chevreul (1786-1889) publicou, aos 53 anos, De la loi du contraste simultané des couleurs (Paris, 1839) que teve sua versão na língua inglesa publicada em 1854. Com essa obra Chevreul estabeleceu, definitivamente, a lei do contraste simultâneo que havia sido mencionada por Leonardo da Vinci (1452-1519) e, posteriormente, por Johann Wolfgang Goethe (1749-1832). Trabalhando desde 1824 na fábrica real de tapetes Manufacture Royal Gobelins (Paris)8, Chevreul teve sua primeira observação do fenômeno do contraste simultâneo com os questionamentos do diretor da fábrica no ano de 1825. O diretor perguntou a Chevreul por que o preto perdia seu vigor quando em contato com sombras circundantes em azul ou violeta (Chevreul, 1854/1987).

Baseado nos seus estudos de contraste e harmonia de cores, Chevreul criou um sistema de cores (Figura 3) para a fábrica que teve como principal contribuição para os próximos sistemas a serem construídos o fato de respeitar os limites de cada matiz, variando as suas distâncias ao centro do sistema. Seu sistema tem como base um círculo cromático com 72 matizes distribuídos radialmente. Para cada matiz pode haver variação da saturação (eixo x), e variação do brilho (eixo y). Os níveis de saturação e brilho podem ter até 22 variações, que variam de acordo com os limites dos matizes (Chevreul, 1854/1987). Esse sistema influenciou diretamente a pintura neo-impressionista de muitos artistas, entre eles Camille Pissaro, Georges Seurat (Figura 4), Paul Signac, Robert Delaunay. Por fim, na Manufacture Royal Gobelins, seu sistema foi utilizado por mais de 150 anos e só foi reorganizado em 1982, devido à introdução de lãs e linhas sintéticas no mercado têxtil (Fischer, 1999; Werner, 1998).

Chevreul foi casado com Sophie Davallet, com quem teve um único filho, Henri, e ficou viúvo aos 76 anos de idade. No seu jubileu centenário, em 1886, o governo francês decidiu reeditar sua obra clássica de 1839. O resultado foi um livro de 586 páginas com as 40 paletas de cores em litografia de alta qualidade, que ainda não era viável na época da primeira edição. Seu filho morreu em 27 de março de 1889. Treze dias depois, em 9 de abril de 1889, Chevreul também morreu. Aos quase 103 anos de idade (Chevreul, 1854/1987).
Ernst Mach (1886-1959), médico, físico e filósofo, além de ser conhecido pela descoberta da velocidade do som, explicou quantitativamente o contraste simultâneo estudado por Chevreul através de apresentações de cartões em preto e branco de forma independente e justaposta: o branco parecia ser mais brilhante e o preto mais escuro em relação à sensação das mesmas quando apresentadas separadamente. Esse fenômeno é conhecido até hoje por Banda de Mach (Mach Band) (Cornsweet, 1970; Kaiser & Boyton, 1996; Backhaus, 1998; Zaidi, 1999).
O mecanismo fisiológico responsável pelo contraste simultâneo é o fenômeno de inibição lateral e foi descrito pelo americano Haldan Keffer Hartline (1903-1983), Prêmio Nobel em Fisiologia ou Medicina (1967). A visão humana é baseada na capacidade de ver bordas, distinguir contrastes entre áreas subjacentes. Se não existissem bordas, não existiria a diferenciação entre os objetos. Hartline demonstrou a inibição lateral em experimentos nos olhos de Limulus, cujos neurônios apresentam dimensões que permitiam registros eletrofisiológicos com maior facilidade. A demonstração foi baseada em registros do efeito de um objeto luminoso sobre o olho do animal em duas situações: em apenas uma face do olho (omatídeo) e em todo olho. No primeiro caso, o resultado é equivalente à medição de um fotômetro. No segundo, há uma intensificação da resposta neural na borda, que confirma o fenômeno de inibição lateral.
Em meados do século XIX (1853), o matemático polonês Hermann Gunter Grassmann (1809-1877) publicou uma teoria da mistura de cores que incluía um conceito fundamental para os futuros sistemas a serem concebidos: a complementaridade das cores, por meio da qual cada ponto em um círculo de cores tem, dentro deste, um complementar (Mollon, 2003).
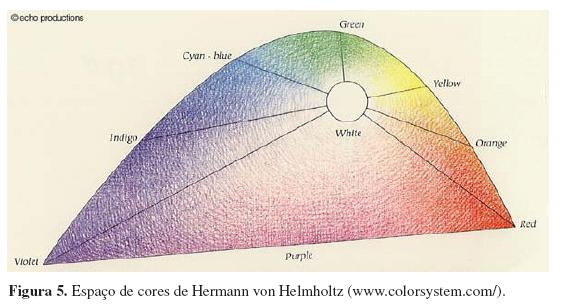
Nesse mesmo período, emergiram os fundamentos básicos para a hipótese de que, uma vez que a luz chega aos olhos, existem três classes de fotorreceptores cones responsáveis pela visão de cores. Essa hipótese foi formulada claramente por Hermann von Helmholtz (1821-1894) e James Clerk Maxwell (1831-1879). Helmholtz, que contribuiu imensamente para a oftalmologia, otologia, neurologia e físico-química, foi o introdutor de um conceito fundamental para os espaços de cores criados no século XX: a demonstração das diferenças nas misturas de cores provenientes de luz (cores-luzes) e de cores provenientes de pigmentos (cores-pigmentos) o que originou a terminologia: mistura aditiva para as cores-luzes e mistura subtrativa para as cores-pigmentos. Helmholtz também é o precursor dos diagramas de cromaticidade. Seu espaço de cores foi considerado o primeiro diagrama de cromaticidade e exerceu forte influência na construção do diagrama de cromaticidade CIE 1931. O espaço de Helmholtz se diferencia dos anteriores também pelo fato de apresentar a gama de púrpuras originada perceptualmente pela combinação das cores situadas nas extremidades do espectro (Figura 5).
Maxwell, por sua vez, foi o responsável pelas primeiras curvas empíricas de misturas de cores-luzes. Seu modelo era baseado em um sistema prismático com 3 fendas, cujas aberturas podiam ser variadas, utilizando várias combinações de comprimentos de onda para a aquisição do branco e, dessa forma, poder obter as curvas de misturas de cores para a reprodução dos comprimentos de onda do vermelho ao violeta. Baseado nessas curvas, Maxwell criou um diagrama de cromaticidade no qual o locus espectral foi determinado pelas proporções das primárias obtidas por seu método psicofísico. Esse método para estimar a sensibilidade espectral dos fotorreceptores da retina foi reproduzido por Helmholtz e Arthur König (1856-1901). O experimento de König, juntamente com Dieterici, foi feito com a participação de indivíduos tricromatas, protanopes e deuteranopes e, a partir desses dados, foram apresentadas as curvas de sensibilidade espectral dos fotorreceptores de um indivíduo tricromata normal: as três curvas de absorção das opsinas dos cones (1886) e a curva de absorção da rodopsina dos bastonetes (1894) (Fisher, 1999; Mollon, 2003).
No final do século XIX, o fisiologista alemão Ewald Hering (1834-1918) desenvolveu a teoria de oponência das cores contrapondo-se a Helmholtz e Maxwell. Com a teoria das oponências, Hering propôs que a experiência da cor resulta da análise, pelo sistema nervoso, das cores em pares opostos. O verde opondo-se ao vermelho. O azul ao amarelo. Assim, explicou o porquê de não sermos capazes de ver “verdes avermelhados” ou “azuis amarelados” e também as pós-imagens negativas que vemos. Mais tarde, adicionou aos canais verde-vermelho e azul-amarelo, o canal branco-preto. Sua teoria de processos oponentes e a teoria tricromática de Young-Newton foram consideradas antagônicas por um longo período. Em um de seus ensaios, Hering indica, em uma nota de rodapé, a co-existência desses dois modelos, a oponência de cores e a tricromacia, no sistema visual humano (Kaiser & Boyton, 1996).
Em conferência informal realizada no dia 5 de fevereiro de 1969, para o Colour Group of Great Britain, W. David Wright (Kaiser & Boyton, 1996), acrescentou algumas contribuições das pesquisas realizadas na área de visão de cores que foram essenciais para a concepção de espaços que possibilitassem a utilização padronizada de medições e especificações de estímulos cromáticos em suas diversas aplicações. Dentre essas contribuições estão as citadas nos parágrafos anteriores desenvolvidas por Grassmannn, Helmholtz, Maxwell, König e Dieterici e, de acordo com Wright, os experimentos, ajustes e deduções das curvas de sensibilidade espectral, realizados nas três primeiras décadas do século XX, pelo físico inglês William Abney, seu colega no Imperial College, e os americanos R. E. Ives e E. A. Weaver.
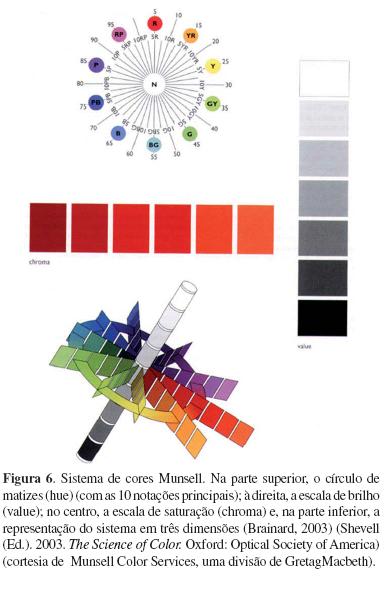
Todas essas descobertas científicas contribuíram para a consolidação dos conceitos básicos da visão de cores e da representação dos mesmos nos espaços a serem concebidos ao longo do século XX. Esses espaços tiveram como referência, em sua grande maioria, o Sistema de Cores Munsell (Figura 6) para os sistemas de aparência de cores e o CIE 1931 (Figura 7) para os sistemas de diferenças de cores.
Alguns anos mais tarde, com a utilização da microespectrofotometria, George Wald (1906-1997) &– Prêmio Nobel em Fisiologia ou Medicina (1967) &– juntamente com Paul K. Brown, apresentaram as curvas de sensibilidade espectral dos fotorreceptores humanos (Brown & Wald, 1964), simultaneamente a Marks, Dobelle e MacNichol (1964) &– em dois artigos da Science. Esses dados impulsionaram diversos cientistas da visão de cores a proporem modificações com a intenção de atualizar os diversos sistemas de cores construídos antes dessa data. Ainda assim, os mais utilizados continuam sendo o Sistema de Cores Munsell (Figura 6) e o CIE 1931 (Figura 7).
Sistema de Cores Munsell
O Sistema Munsell (Figura 6) foi desenvolvido pelo artista e educador Albert Henry Munsell (1858-1918), tendo sido publicado como um sistema de ordenamento de cores em 1905. Nesse sistema, as cores são arranjadas cilindricamente com as coordenadas descritas a seguir: o eixo vertical representa value (V), uma dimensão associada ao brilho, entre preto (V = 0) e branco (V = 10), para as cores localizadas respectivamente nos extremos &– inferior e superior &– do eixo vertical. Perpendicularmente ao eixo vertical estão dispostas as outras dimensões representando os outros dois atributos das cores. O ângulo polar e a distância, representam, respectivamente, o hue ou matiz (H) e o chroma (C), dimensão associada à saturação. No caso do matiz, o círculo de cores do sistema é dividido em 100 partes iguais, identificadas no círculo externo, H variando de 0 a 100. Por exemplo, os matizes vermelhos (R), encontram-se entre 0 ≤ H ≤ 10. A saturação, determinada pelo afastamento em relação ao eixo vertical, mostra que uma cor, quanto mais distante do eixo vertical, maior o seu grau de saturação. Quando foi elaborado, esse sistema propunha que as cores fossem arranjadas de forma que duas cores vizinhas tivessem a mesma magnitude de diferença perceptual. Tal finalidade não foi alcançada pelo sistema, mas sua eficiência na especificação de cores é comprovada devido ao fato de continuar sendo utilizada por diversas indústrias ao longo de mais de cem anos. Suas especificações também são utilizadas na elaboração de testes psicofísicos para avaliação de deficiências na visão de cores. A versão atual, a Renotation Munsell, é baseada em um espaço visual um pouco mais abrangente, contendo 1928 pontos. Essa versão foi aprimorada e adotada pelo Comitê de Colorimetria da Optical Society of America (Birch, 2001; Brainard, 2003; Indow, 1995).
CIE 1931
A colorimetria tornou-se tecnicamente viável quando foram apresentadas as Recommendations officielles de la Comission Internationale de l'Eclairage (1931) (Schanda, 1998) determinadas pela sistematização das funções de misturas de cores necessárias para um observador padrão dentro de um campo visual de 2o, em condições específicas de iluminação.
A retina humana possui três tipos de cones, com sensibilidade máxima em diferentes regiões do espectro visível, mas com grande sobreposição entre as faixas espectrais de absorção de luz. As diferentes misturas das três primárias possibilitam a formação de todas as cores possíveis de serem percebidas pelo homem. Dessa forma, as cores podem ser obtidas quando se têm as proporções adequadas para cada primária. Essa mistura ocorre quando cores-luzes coincidem no espaço e no tempo de modo a serem vistas como uma única cor-luz (Rodrigues, 2003).
A construção do espaço de cores CIE foi realizada com a utilização da equiparação cromática. Nessa técnica, é apresentado ao observador um campo circular bipartido, com um dos lados iluminado por uma luz monocromática e a outra metade sendo iluminada simultaneamente por três diferentes luzes que, em geral, correspondem às primárias vermelho, verde e azul. A tarefa do observador é ajustar a intensidade de cada uma das três primárias até que as duas metades do círculo sejam percebidas como iguais. A trinca de valores passa a representar a cor monocromática que foi testada. O experimento é realizado em intervalos de aproximadamente 4 nm (Rodrigues, 2003).
O sistema de cores CIE 1931 (XYZ) é conhecido pelo seu diagrama de cromaticidade (Figura 7). A forma tridimensional do sistema pode ser obtida calculando-se as coordenadas do terceiro eixo (Y), o eixo de luminância, somadas às coordenadas dos dois eixos de cromaticidade (x e y).
No diagrama estão representadas não apenas as cores puras ou monocromáticas como, também, as misturas delas para cada coordenada cromática. As coordenadas para as cores puras são representadas no locus espectral, o qual ocupa mais de dois terços da borda do diagrama, correspondendo aos comprimentos de onda entre 380 nm e 780 nm. O branco, por sua vez, está localizado em sua porção mais central (x = 0,33; y = 0,33) e a sua determinação resulta da combinação dos três comprimentos de onda adotados como primárias, 700 nm, 546,1 nm e 435,8 nm. Esses valores foram obtidos por Guild (1931, citado por Wright, 1996/1998) em um de seus experimentos e estão bem próximos dos encontrados por Wright (1929) (Wright, 1969/1998; Simões, 2004).
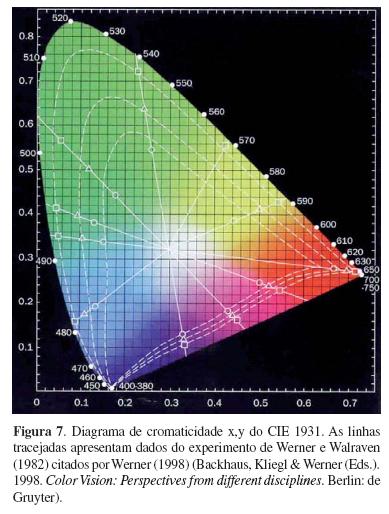
No sistema, X, Y e Z representam cada um dos valores absolutos da mistura de cores das três primárias necessárias para a equiparação com cada uma das cores espectrais. Os valores de x, y e z representam as coordenadas de cromaticidade e são obtidas pelas fórmulas: x = X/(X+Y+Z), y = Y/(X+Y+Z) e z = Z/(X+Y+Z). Na representação gráfica do diagrama, as informações são representadas em x, y e Y. As coordenadas x e y são os coeficientes correspondentes às quantidades relativas de dois das três primárias e a da terceira, z, é obtida por diferença, uma vez que x + y + z = 1, ou seja, z = 1 &– x &– y. Estas três coordenadas correspondem às proporções de cada uma das três primárias para constituir cada cor espectral. Desta forma, as cores têm informações nas três dimensões, enquanto o branco e o preto estão apenas no eixo Y que representa o nível de luminância expresso em cd/m2 (Kaiser & Boynton, 1996).
O CIE 1931 vem sendo utilizado sem alterações substanciais desde a sua concepção. Alguns pontos foram interpolados ou aprimorados, seus valores numéricos foram expressos com aproximação de sete dígitos, mas os fundamentos continuam os mesmos (Schanda, 1998).
Há muitas limitações que hoje são mais conhecidas. Uma delas é a situação hipotética de iluminação que não representa as situações cotidianas de percepção das cores. Outra limitação é o fato de não representar fielmente o sistema da visão de cores humana (Kaiser & Boynton, 1996). Como não se tem, ainda, um sistema que reproduza fielmente a percepção de cores humana e sua respectiva representação, o diagrama de cromaticidade CIE 1931 e suas modificações ainda são muito utilizados em métodos psicofísicos de avaliação da visão de cores.
Outros sistemas de cores
Existem vários outros sistemas de cores, sejam de aparência de cores ou de diferenças: o Swedish Natural Colour System (NCS), o OSA Uniform Color Scale (OSA/UCS), o DIN Color System (DIN), RGB, Gerritsen, MacLeod-Boyton, DKL, CIELAB, CIE Rösch, CIE MacAdam, CIE Walter Stiles, CIE 1976, entre outros (Backhaus, Kliegl, & Werner, 1998; Gerritsen, 1974; Kaiser & Boyton, 1996; Shevell, 2003).
A reconstrução dos espaços de cores
As propriedades físicas dos estímulos cromáticos são especificadas pela composição espectral do estímulo e pela intensidade medida em unidades de energia ou número de quanta. A composição espectral é traduzida nos sistemas de representação de cores em coordenadas tricromáticas que correspondem ao matiz, à saturação e ao brilho. O brilho corresponde à intensidade. Matiz, saturação e brilho são atributos psicológicos, que são influenciados não apenas pelos atributos físicos do estímulo, como também por vários outros fatores como, por exemplo, pelos demais estímulos presentes no campo visual. A proximidade de outros estímulos modifica a percepção de cor por efeitos como indução cromática (Zaidi, 1999), assimilação (Kaiser & Boyton, 1996; Werner, 1998) e contraste simultâneo (Chevreul, 1839/1987; Kaiser & Boyton, 1996). O sistema visual também exibe constância de cor, ou seja, a despeito de mudanças grandes no iluminante, atribui-se a mesma cor a um objeto, o que mostra que o atributo físico e a refletância espectral não determinam percepção cromática de modo absoluto e que a percepção da cor é influenciada pelo contexto. Brainard (2003) defende a idéia de que, para várias aplicações, o que importa é saber a aparência cromática do estímulo e ter uma forma fácil de, a partir dessa especificação, chegar às coordenadas de cromaticidade que o especificam fisicamente.
Um outro aspecto de extrema importância na especificação dos estímulos cromáticos é a magnitude da diferença entre os estímulos. Na construção de espaços de cores que visam reproduzir as diferenças mínimas entre os estímulos cromáticos, encontra-se uma pergunta crucial: quão diferentes devem ser as coordenadas de matiz, brilho e saturação de dois estímulos de cores para reproduzir a mínima diferença perceptível? (Brainard, 2003).
Essa dificuldade em distinguir os aspectos físicos e perceptuais da cor pode ser exemplificada por um dos fenômenos de contraste simultâneo, o efeito de assimilação. Nesse fenômeno, um painel com o fundo em uma mesma cor, quando intercalado por uma grade preta, a cor aparenta ser mais escura e, quando intercalada pela grade branca, aparenta ser mais clara. Tal efeito demonstra a diferença na sensação da mesma cor em diferentes contextos (Backhaus, 1998; Werner, 1998).
Para que a relação entre as percepções de cores seja representada em sua totalidade, é preciso conhecer completamente os mecanismos fisiológicos do sistema visual, assim como representar a magnitude das diferenças entre elas. A compreensão dessas relações possibilita a melhor representação das mesmas e permite conceber espaços perceptuais de cores que mais se aproximam da realidade perceptual. Alguns autores referem-se a esse processo como uma reconstrução, uma vez que se estaria procurando recriar o espaço perceptual que cada indivíduo possui para representar as cores (e.g.: Paramei, 1996; Paramei, Bimler, & Cavonius, 2001).
Os espaços de cores, como dito anteriormente, podem ser obtidos a partir de dados fisiológicos ou psicofísicos. Cornsweet (1970) construiu um espaço de cores baseado na capacidade de absorção de luz dos cones da retina humana, no qual cada eixo era constituído pela curva de absorção de um dos fotopigmentos. Na psicofisiologia comparada da visão de cores, esse tipo de espaço é obtido com métodos intracelulares eletrofisiológicos de determinação da sensibilidade espectral de cada fotorreceptor, resultando num espaço cromático que considera a entrada espectral do sistema visual e que permite a localização de objetos relevantes para a espécie. Dessa forma, pode-se examinar se as distâncias nesse espaço correspondem a diferenças de discriminação cromática. Os espaços de cores construídos para invertebrados (Menzel, 1979; Menzel, Ventura, Hertel, Souza, & Greggers, 1986; Peitsch et al., 1992) e vertebrados (Neumeyer, 1984; Neumeyer & Arnold, 1989), serviram para comparar dados de entrada espectral com os dados de discriminação de cores nesses animais, o que permitiu testar hipóteses sobre os mecanismos responsáveis pela discriminação de cores (Backhaus, 1998).
No caso específico das abelhas, foram realizados estudos fisiológicos e psicofísicos detalhados para a obtenção de seus espaços de cores. Frisch (1914, citado por Backhaus, 1998) demonstrou a capacidade das abelhas em discriminar um azul de uma série de cinza. Daumier (1956, citado por Backhaus, 1998) confirmou a tricromacia da visão de cores das abelhas através das apresentações de luzes indistinguíveis de uma referência, chamadas cores metaméricas, determinadas por diversas combinações de luzes monocromáticas primárias especificadas num determinado diagrama de cromaticidade, o que resultou em uma reconstrução tridimensional do espaço de cores. O primeiro experimento eletrofisiológico foi realizado por Autrum e Zwehl (1964, citado por Backhaus, 1998) para medir a sensibilidade espectral dos três fotorreceptores das abelhas e, mais tarde com maior precisão, por Menzel et al. (1986) ao testar espécies tropicais de abelhas.
O conhecimento da função de sensibilidade espectral dos fotorreceptores das abelhas possibilitou a obtenção de seus espaços de cores por Helversen (1972, citado por Backhaus, 1998) e Backhaus e Menzel (1987, citado por Bachaus, 1998) e a concepção de outros espaços que associaram essas informações ao conceito das mínimas diferenças perceptíveis das cores por Backhaus (1998). Nesse experimento, os resultados demonstraram a existência do efeito de Bezold-Brucke (Cornsweet, 1970; Kaiser & Boyton, 1996) para as abelhas, equivalente ao já conhecido para a visão humana. Foram utilizadas técnicas intracelulares para a determinação da atividade neuronal dos canais de oponência por Kien e Menzel (1977, citado por Backhaus, 1998). Experimentos nesse sentido, como os de Helversen (1972, citado por Backhaus, 1998) e Brandt et al. (1993, citado por Backhaus, 1998), mostraram que os valores de excitação dos neurônios dos canais de oponência de cor correspondem às coordenadas obtidas pelos resultados psicofísicos de discriminação de cores das abelhas.
Uma das formas utilizadas para a reconstrução de espaços de cores é colher dados de julgamentos de semelhanças ou diferenças entre os estímulos de cores. A determinação “métrica” da relação entre os dados é gerada, normalmente, pela técnica de escalonamento multidimensional, que reconstrói o espaço de cores do sujeito a partir do conjunto de distâncias relativas. Esse procedimento é mais comumente conhecido como escala multidimensional (mds), que pode ser métrica (espaço Euclideano) ou não-métrica (espaço Minkowski) (Backhaus, 1998; Kruskal & Wish, 1978). O primeiro procedimento métrico foi realizado por Richardson (1938, citado por Kruskal & Wish, 1978), baseado no teorema de Young e Householder (1938), muito próximo da análise fatorial, na qual a configuração das relações é dada pelas distâncias Euclideanas entre os estímulos. Esse método foi mais tarde aprimorado por Torgerson (1958, citado por Kruskal & Wish, 1978). No método não-métrico, as relações entre os estímulos não são dependentes das diferenças ou atributos já conhecidos entre os pontos analisados e, por isso, dá-se o nome não-métrico. O primeiro procedimento não-métrico foi descrito por Coombs (1964, citado por Kruskal & Wish, 1978), no qual a configuração das relações é obtida exclusivamente pelos julgamentos de diferenças e semelhanças através de funções monotônicas crescentes ou decrescentes para a representação das relações em duas ou três dimensões. O interesse de Boyden (1933, citado por Kruskal & Wish, 1978) pela aplicação biológica do mds foi registrado historicamente pelo estudo da proximidade entre as espécies determinada pela intensidade da reação entre corpos e anticorpos. Outras formas de análise podem ser obtidas, por exemplo, pela derivação de soluções métricas a partir da mds não-métrica que foi particularmente estudada por Shepard (1962).
Atualmente, a mds é utilizada nas mais diversas áreas como, por exemplo, a reconstrução de árvores filogenéticas de organismos, ou o estudo da discriminação de produtos em marketing. Essa técnica estatística multivariada passou a ser mais utilizada somente a partir da década de 80, com o desenvolvimento da microeletrônica e a conseqüente redução dos custos computacionais. Os cálculos necessários à mds são elaborados e mesmo as versões mais simples necessitam de programas para a análise dos dados (Davison & Sireci, 2000; Helm, 1964; Indow, 1995; Kruskal & Wish, 1978; Shepard, 1962; Shepard & Cooper, 1992).
A dificuldade na sistematização dos espaços de diferenças de cores está na reprodução de todas as diferenças minimamente perceptíveis existentes entre os estímulos de cores. Pesquisadores de diversas áreas, dentre eles psicofísicos e neurofisiologistas, continuam estudando os caminhos pelos quais se possa construir um espaço de cores capaz de abranger as diversas variações perceptuais da visão de cores, seja humana ou animal (Helm & Tucker, 1962; Paramei & Bimler, 2001b). Para tal, as unidades métricas de seus sistemas devem estar associadas às magnitudes de suas diferenças: a mesma distância entre quaisquer dos dois pontos representaria igual capacidade de discriminação ou igual diferença perceptual. Esse tipo de espaço recebe o nome, por alguns autores, de espaço uniforme de cores (Helm & Tucker, 1962; Paramei & Bimler, 2001b; Paramei, Izmailov, & Sokolov, 1991; Thompson, Palácios, & Varela, 1992).
Nos últimos 25 anos, muitos trabalhos se dedicaram à reconstrução de espaços de cores de indivíduos normais e de indivíduos com deficiências congênitas (Bimler, 1999; Bimler & Kirkland, 2002; Bimler, Kirkland, & Jacobs, 2000; Bimler & Paramei, 2004; Izmailov & Sokolov, 1991; Paramei et al., 1991; Paramei, 1996; Paramei, Bimler, & Cavonius, 1998; Paramei & Bimler, 2001a, 2001b; Paramei, Bimler, & Cavonius, 2001). Muitos desses trabalhos propuseram a visualização do espaço deficitário como uma deformação do espaço normal. Embora de extrema importância para o conhecimento sobre o sistema visual humano, esses trabalhos ainda não podem ter aplicação clínica, pois reportam a reconstrução do espaço de cores para um conjunto de indivíduos.
Feitosa-Santana (2005) partiu desses trabalhos, estendendo a proposta para a reconstrução de espaços de cores de pacientes com deficiência adquirida na visão de cores e que puderam ser comparados aos espaços de cores de sujeitos normais. Os testes de visão de cores (D-15 e D-15d) em que fichas de diferentes cores são ordenadas pelo indivíduo testado, foram aplicados em procedimento tradicional &– para diagnóstico da visão de cores &– e em procedimento triádico &– para a reconstrução dos espaços de cores. No procedimento triádico, as fichas foram apresentadas em trios e o sujeito deveria separar a que mais diferisse (odd-one-out). Os testes foram aplicados em dois grupos experimentais: pacientes com diabetes mellitus tipo 2 sem nenhum sinal de retinopatia (Feitosa-Santana, Oiwa et al., 2006) e ex-trabalhadores de fábricas de lâmpadas fluorescentes intoxicados por vapor de mercúrio (Feitosa-Santana, Paramei et al., 2006); e seus respectivos grupos controles. Os espaços de cores foram reconstruídos para os indivíduos e os grupos e, representados em forma bi-dimensional, onde as dimensões correspondem aos eixos verde-vermelho (RG) e azul-amarelo (BY) dos sistemas de oponência perceptual. No procedimento tradicional, as perdas dos grupos experimentais foram predominantes no eixo BY e, na reconstrução dos espaços de cores, as perdas se apresentaram em ambos os eixos, RG e BY, classificadas como difusas. Os resultados confirmam os achados de perdas difusas na visão de cores de pacientes diabéticos tipo 2 sem retinopatia (Gualtieri, 2004; Santana et al., 2005; Ventura et al., 2003) e intoxicados por vapor de mercúrio (Feitosa-Santana, Costa, Lago, & Ventura, 2007; Silveira et al. 2003; Silveira, Ventura, & Pinheiro, 2004; Ventura et al. 2004; Ventura et al., 2005). Esse trabalho concluiu que os espaços de cores dos grupos de pacientes com diabetes mellitus tipo 2 sem retinopatia (Feitosa-Santana et al., 2006) e de pacientes intoxicados por vapor de mercúrio (Feitosa-Santana et al., 2006) diferem dos espaços de cores dos respectivos grupos controle.
O estudo ainda sugere que a análise quantitativa da reconstrução dos espaços de cores, por ser uma medida objetiva da percepção do sujeito, pode ser eficiente na distinção entre sujeitos com deficiência e sujeitos normais. A adoção de tal método para as análises individuais, além de pouco dispendioso, pode ter impacto nos testes de diagnóstico ou acompanhamento clínico de diversas doenças que afetam o sistema visual, tais como: a diabetes, a doença de Parkinson, a esclerose múltipla, as intoxicações por metais pesados e outras.
Referências
Backhaus, W. G. K. (1998). Physiological and psychological simulations of color vision in humans and animals. In W. G. K. Backhaus, R. Kliegl & J. S. Werner (Eds.), Color vision: Perspectives from different disciplines, (pp. 45-78). Berlin: de Gruyter. [ Links ]
Backhaus, W. G. K., Kliegl, R., & Werner, J. S. (Eds.). (1998). Color vision: Perspectives from different disciplines. Berlin: de Gruyter. [ Links ]
Bimler, D. (1999). A multidimensional scaling comparison of color metrics for response times and rated dissimilarities. Perception & Psychophysics, 61, 1675-1680. [ Links ]
Bimler, D., Kirkland, J., & Jacobs, R. (2000). Colour-vision tests considered as a special case of multidimensional scaling. Color Research and Application, 25, 160-169. [ Links ]
Bimler, D. & Kirkland, J. (2002). Sex differences in color vision and the salience of color-space axes. Journal of Vision, 2, 28a. [ Links ]
Bimler, D. & Kirkland, J. (2004). Multidimensional scaling of D15 caps: Color vision defects among tobacco smokers? Visual Neuroscience, 21, 445-448. [ Links ]
Bimler, D. & Paramei, G. (2004). Luminance-dependent hue shift in protanopes. Visual Neuroscience, 21, 403-407. [ Links ]
Birch, J. (2001). Diagnosis of defective colour vision. Oxford: Butterworth Heinmann. [ Links ]
Brainard, D. (2003). Color appearance and color difference specification. In S. Shevell (Ed.), The science of color, (pp. 191-216). Oxford: Optical Society of America. [ Links ]
Brown, P. K. & Wald, G. (1964). Visual pigments in single rods and cones of the human retina. Science, 144(3614), 45-52. [ Links ]
Campenhausen, C. A. (2001). Preliminary form of 3-dimensional color systems in Rubens' illustrated textbook of Franciscus Aguilonius (1613). Medizinhistorisches Journal, 36, 267-307. [ Links ]
Chevreul, M. E. (1987). The principles of harmony and contrasto f colors and their applications to the arts (A newly revised edition with special introduction and newly revised commentary by Faber Birren). West Chester, PA: Schiffer. (Trabalho original publicado em 1839) [ Links ]
Cornsweet, T. (1970). Visual perception. Oxford: Academic Press. [ Links ]
Davison, M. & Sireci, S. (2000). Multidimensional scaling. In H. E. A. Tinsley, S. D. Brown (Eds.), Handbook of applied multivariate statistics and mathematical modeling. San Diego, CA: Academic Press. [ Links ]
De Valois, R. L., Abramov, I., & Mead. W. R. (1967). Single cell analysis of wavelength discrimination at the lateral geniculate nucleus in the macaque. Journal Neurophysiology. 30(3), 415-433. [ Links ]
Feitosa-Santana, C. (2005). Reconstrução do espaço de cores de pacientes com discromatopsia adquirida: diabéticos tipos 2 e intoxicados por vapor de mercúrio. Dissertação de Mestrado, Instituto de Psicologia, Universidade de São Paulo, São Paulo. [ Links ]
Feitosa-Santana, C., Oiwa, N., Paramei, G., Bimler, D., Costa, M. F., Lago, M. et al. (2006). Color space distortions in patients with type 2 diabetes mellitus. Visual Neuroscience, 23, 663-668. [ Links ]
Feitosa-Santana, C., Paramei, G., Bimler, D., Costa, M. F., Lago, Oiwa, N. et al. (2006). Color space distortions in patients with vapor mercury intoxication. In European Conference on Visual Perception, 27, St. Petersburg. Perception, 35(Suppl.), 185-186. [ Links ]
Feitosa-Santana, C., Costa, M. F., Lago, M., & Ventura, D. F. (2007). Loss of long-term color vision after exposure to mercury-vapor. Brazilian Journal of Medical and Biological Research, 40(3), 409-414. [ Links ]
Fischer, E. (1999). Colour order systems in art and science. Recuperado em 10 de dezembro de 2005 e 5 de janeiro de 2006, de http://www.colorsystem.com [ Links ]
Gage, J. (2001). Colour and culture: Practice and meaning from antiquity to abstraction. London: Thames & Hudson. [ Links ]
Gerritsen, F. (1974). Présence de la couleur. Paris: Dessain et Tolra. [ Links ]
Gualieri, M. (2004). Visão de cores e sensibilidade ao contraste em indivíduos com diabete melito: avaliação psicofísica e eletrofisiológica. Dissertação de Mestrado, Instituto de Psicologia, Universidade de São Paulo, São Paulo. [ Links ]
Helm, C. (1964). Multidimensional ratio scaling analysis of perceived color relations. Journal of the Optical Society of America, 54, 256-262. [ Links ]
Helm, C. & Tucker, L. (1962). Individual differences in the structure of color-perception. American Journal of Psychology, 75, 437-444. [ Links ]
Indow, T. (1995). Psychophysical scaling: Scientific and practical applications. In R. D. Luce, M. D. D'Zmura, D. Hoffman, G. J. Iverson & A. K. Romney (Eds.), Geometric representations of perceptual phenomena (pp. 1-34). Mahwah, NJ: Erlbaum. [ Links ]
Izmailov, C. & Sokolov, E. (1991). Spherical model of color and brightness discrimination. Psychological Science: a journal of the American Psychological Society / APS 2, 249-259. [ Links ]
Jaeger, W. (1984). Principles of order in the color systems of the 17th century. Klinische Monatsblätter für Augenheilkunde., 184(4), 321-325. [ Links ]
Kaiser, P. K. & Boynton, R. M. (1996). Human color vision. Washington DC, USA: Optical Society of America. [ Links ]
Kruskal, J. B. & Wish, M. (1978). Multidimensional scaling. In M. S. Lewis-Beck (Ed.), Quantitative applications in the Social Sciences. Londres: Sage. [ Links ]
Kuehni, R. & Stanziola, R. (2002). Francis Glisson's color specification system of 1677. Color Research and Application, 27, 15-19. [ Links ]
Marks, W. B., Dobelle, W. H., & MacNichol Jr., E. F. (1964). Visual pigments of single primate cones. Science, 143(3611), 1181-1182. [ Links ]
Menzel, R. (1979). Spectral sensitivity and color vision in invertebrates. In H. Autrum (Eds.), Comparative physiology and evolution of vision in invertebrates. Handbook of sensory physiology (Vol. 7, p. 6A). Berlin: Springer. [ Links ]
Menzel, R., Ventura, D. F., Hertel, H., Souza, J. M., & Greggers, U. (1986). Spectral sensitivity of photoreceptors in insect compound eyes: Comparison of species and methods. Journal of Comparative Physiology, 158, 165-177. [ Links ]
Mollon, J. (2003). The origins of modern color science. In S. Shevell (Ed.), The science of color. (pp. 1-39). Oxford: Optical Society of America. [ Links ]
Neumeyer, C. (1984). On spectral sensitivity in the goldfish. Evidence for neural interactions between different “cone mechanisms”. Vision Research, 24, 1223-1231.
Neumeyer, C. & Arnold, K. (1989). Tetrachromatic color vision in the goldfish becomes trichromatic under white adaptation light of moderate intensity. Vision Research, 29, 1719-1727. [ Links ]
Paramei, G. (1996). Color space of normally sighted and color-deficient observers reconstructed from color naming. Psychological Science, 7, 311-317. [ Links ]
Paramei, G. V. & Bimler, D. L. (2001a). Vector coding underlying individual transformations of a color space. In C. Musio (Ed.), Vision. The approach of biophysics and neurosciences (pp. 429-436). New Jersey: World Scientific. (Proc. of the International School of Biophysics, Series on Biophysics and Biocybernetics, 11. Casamicciola, Napoli, Italy, 11-16 Oct. 1999) [ Links ]
Paramei, G. V. & Bimler, D. L., (2001b). Is color space curved? A common model for color-normal and color-deficient observers. In W. G. K. Backhaus (Ed.), Neuronal coding of perceptual systems (pp. 102-105). New Jersey: World Scientific. (Proc. of the International School of Biophysics, Series on Biophysics and Biocybernetics, 9. Casamicciola, Napoli, Italy, 12-17 Oct. 1998) [ Links ]
Paramei, G. V., Bimler, D. L., & Cavonius, R. (1998). Effect of luminance on color perception of protanopes. Vision Research, 38, 3397-3401. [ Links ]
Paramei, G. V., Bimler, D. L., & Cavonius, C. R. (2001). Color-vision variations represented in an individual-difference vector chart. Color Research and Application, 26(Suppl.), 230-234. [ Links ]
Paramei, G., Izmailov, C., & Sokolov, E. (1991). Multidimensional scaling of large color differences. Psychological Science, 2, 244-248. [ Links ]
Paramei, G. (2004). Singing the russian blues: An argument for culturally basic color terms. In Biennial Conference Cross-Cultural Research, 7, Workshop “Anthropology of colour: Colour as a phenomenon of culture”, 39, 10-34.
Parkhurst, C. & Feller, R. L. (1982). Who invented the color wheel? Color Research and Application, 7, 217-230. [ Links ]
Peitsch, D., Fietz, A., Hertel, H., Souza, J., Ventura , D. F., & Menzel, R. (1992). The spectral input systems of hymenopteran insects and their receptor-based colour vision. Journal of Comparative Physiology A, 170, 23-40. [ Links ]
Rodrigues, A. R. (2003). O desenvolvimento de aplicativos de avaliação psicofísica visual em estações de trabalho IBM RISC 6000. Dissertação de Mestrado, Instituto de Ciências Biológicas, Universidade Federal do Pará, Belém. [ Links ]
Santana, C. F., Costa, M. F., Lago, M., Bernik, M., Nishi, M., & Ventura, D. F. (2005). Color discrimination in type 2 diabetes mellitus patients with no retinopathy. Investigative Ophthalmology & Vision Sciences, 46, E-Abstract 4579. (ARVO. Annual Meeting (Association for Research in Vision and Ophthalmology), Ft. Lauderdale. Rockville, MD: Association for Research in Vision and Ophthalmology) [ Links ]
Schanda, J. (1998). Current CIE work to achieve physiologically-correct color metrices. In W. G. K. Backhaus, R. Kliegl, & J. S. Werner (Eds.), Color vision: Perspectives from different disciplines (pp. 307-318). Berlin: de Gruyter. [ Links ]
Shepard, R. N. (1962). The analysis of proximities: Multidimensional scaling with an unknown distance function. Psychometrika, 27, 125-140, 219-246. [ Links ]
Shepard, R. N. & Carroll, J. D. (1966). Parametric representation of nonlinear data structures. In P. R. Krishnaiah (Ed.), Multivariate analysis (pp. 561-592). Dayton, OH: Academic Press. (Proceedings of an International Symposium, 14-19 jun. 1965) [ Links ]
Shepard, R. N. & Cooper, L. A. (1992). Representation of colors in the blind, color-blind, and normally sighted. Psychological Science, 3, 97-104. [ Links ]
Shevell, S. (Ed.). (2003). The science of color. Oxford: Optical Society of America. [ Links ]
Silveira, L. C. L., Damin, E. T. B., Pinheiro, M. C. N., Rodrigues, A. R., Moura, A. L. A., Cortês, M. I. T., & Mello, G. A. (2003). Visual dysfuntion following mercury exposure by breathing mercury vapor or by eating mercury contaniminated food. In J. D. Mollon, J. Pokorny & K. Knoblauch (Eds.), Normal and defective color vision (pp. 409-417). London: Oxford Press. [ Links ]
Silveira, L. C. L., Ventura, D. F., & Pinheiro, M. C. N. (2004). Toxidade mercurial: avaliação do sistema visual em indivíduos expostos a níveis tóxicos de mercúrio. Ciência e Cultura, 56(1), 36-38. [ Links ]
Simões, A. L. A. C. (2004). Estudo das alterações visuais cromáticas em trabalhadores expostos ao vapor de mercúrio. Dissertação de Mestrado, Instituto de Psicologia, Universidade de São Paulo, São Paulo. [ Links ]
Thompson, E., Palacios, A., & Varela, F. (1992). Ways of coloring: Comparative color vision as a case of study for cognitive science. Behavioral and Brain Sciences, 15, 1-74. [ Links ]
Ventura, D. F., Costa, M. F., Gualtieri, M., Nishi, M., Bernik, M, Bonci, M. et al. (2003). Early vision loss in diabetics patients assessed by the Cambridge Colour Test. In J. D. Mollon, J. Pokorny & K. Knoblauch (Eds.), Normal and defective color vision (pp. 391-399). London: Oxford Press. [ Links ]
Ventura, D. F., Costa, M. T. V., Costa, M. F., Berezovsky, A., Salomão, S. R., Canto-Pereira, L. H. M. et al. (2004). Multifocal and full field electroretinogram changes associated with color vision loss in mercury vapor exposure. Visual Neuroscience, 21, 421-429. [ Links ]
Ventura, D. F., Simões, A. L., Canto-Pereira, L. H. M., Tomaz, S., Lago, M., Costa, M. T. V. et al. (2005). Color vision and contrast sensitivity losses of mercury contaminated industry workers in Brazil. Environmental Toxicology and Pharmacology, 19, 523-529. [ Links ]
Werner, J. S. (1998). Aging through the eyes of Monet. In W. G. K. Backhaus, R. Kliegl, & J. S. Werner (Eds.), Color vision: Perspectives from different disciplines (pp. 3-41). Berlin: de Gruyter. [ Links ]
Wright, W. D. (1969/1998). Part II: The origins of the 1931 CIE System. In P. K. Kayser & R. M. Boynton (Eds.), Human color vision (Appendix, Part II, pp. 534-543). Washington, DC: Optical Society of America. [ Links ]
Zaidi, Q. (1999). Color and brightness induction: From mach bands to three dimensional configurations. In K. R. Gegenfurtner & L. T. Sharpe, Color vision: From genes to perception, 317-344. Cambridge, UK: Cambridge University Press. [ Links ]
Recebido em: 19/11/2006
Aceito em: 8/12/2006
1 Artigo originado da dissertação de mestrado da primeira autora defendida em 2006 no Instituto de Psicologia - USP. Apoio financeiro do Projeto Temático FAPESP, CNPq e CAPES/PROCAD. Agradecimento especial a revisão de Lourdes Ortellado.
2 Pós-Graduanda de Neurociências e Comportamento do Instituto de Psicologia
- USP. Endereço eletrônico: claudia@feitosa-santana.com
3 Colaborador do Laboratório da Visão coordenado pela Profa. Titular Dora Fix Ventura.
4 Docentes do Departamento de Psicologia Experimental do Instituto de Psicologia - USP.
5 Docente do Departamento de Fisiologia do Centro de Ciências Biológicas - UFPA. Diretor Geral do Núcleo de Medicina Tropical - UFPA.
6 John Mollon (8 de julho, 2005) no 18th Symposium of the International Color Vision Society (Lyon, Paris), homenageando Monge, distribuiu folhas de acetato na cor vermelha para que todos ali presentes pudessem repetir a experiência.
7 De acordo com Mollon (2003), o fenômeno das Sombras Coloridas foi relatado anteriormente, em 1672, por Otto Von Guericke (1602-1686), o inventor da bomba a vácuo.
8 Chevreul foi nomeado para o cargo de Diretor Colorista da Manufacture Royal Gobelins no dia 9 de setembro de 1824, pelo Rei Luís XVIII. Até hoje a fábrica é controlada pelo governo, mas no dia 13 de julho de 2005, um de nossos autores esteve lá para uma visita e presenciou uma triste manifestação dos funcionários da fábrica contra o possível fechamento da mesma devido a não lucratividade e não valorização da arte realizada nessa fábrica em atividade há centenas de anos. A sociedade moderna, em cujos altares cultua-se o Lucro, um deus que substituiu todos os demais, indoarianos ou não, surpreende-nos cotidianamente e além de qualquer medida pela sua insensatez.
