








Servicios Personalizados
Articulo
 Portugués (pdf)
Portugués (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
Indicadores

Compartir

 Permalink
PermalinkArquivos Brasileiros de Psicologia
versión On-line ISSN 1809-5267
Arq. bras. psicol. vol.62 no.1 Rio de Janeiro abr. 2010
ARTIGOS
Tendências em arquiteturas cognitivas
Trends in cognitive architectures
Álvaro Machado Dias
Universidade de São Paulo, São Paulo, Brasil
RESUMO
Ao longo do último século, modelos voltados a revelar como a mente emerge do substrato orgânico levaram ao desenvolvimento de arquiteturas cognitivas. Atualmente, esta tendência se encontra em franco desenvolvimento, por meio da introdução de novos e decisivos princípios. Este artigo introduz uma revisão histórica deste campo, seguida pela apresentação do mais célebre desses novos princípios, o de “redes de pequenos mundos”. Em seguida, consideramos uma revisão sistemática da distribuição dos estudos em “redes cognitivas de pequenos mundos” (2007-2009). Por fim, os novos conceitos em arquiteturas cognitivas são aplicados para a compreensão etiológica e nosológica da esquizofrenia.
Palavras-chave: Cognição; Relações mente-corpo; Arquitetura Cognitiva; Psicopatologia.
ABSTRACT
In the last century, models addressing the emergence of mind from its organic basis led to the development of Cognitive Architectures. Nowadays, this tendency is facing great development, by means of the introduction of original and crucial principles. This paper introduces a historical review of this field, followed by the introduction of the most remarkable of these new principles, “small world networks”. Next, the paper proceeds with a systematic review of recent studies in “small world cognitive networks” (2007-2009). At last, the new concepts in cognitive architecture are applied for the etiological and nosological comprehension of schizophrenia, psychopathology’s distinguished puzzle.
Keywords: Cognition; Mind-body relations; Cognitive Architecture; Psychopathology.
INTRODUÇÃO
Materialismo como Pedra de Toque nos Estudos sobre o Funcionamento da Mente
O presente artigo visa introduzir o conceito de arquitetura neural e alguns de seus principais usos na ciência cognitiva desenvolvida na atualidade.
Alinhavando máximas científico-filosóficas tão festejadas no âmbito das ciências naturais quanto o darwinismo neural (para uma revisão, ver Edelman (1993)) e o princípio da computabilidade/incomputabilidade em sistemas biológicos (DAVIS, 1965), o princípio de que o funcionamento da mente é correlato ao comportamento das células nervosas ou, em outras palavras, a inexorabilidade da condição mente/sistema nervoso é o ponto de partida para a imensa maioria dos estudos atuais em ciência cognitiva (para uma revisão, ver Churchland (1986)).
Tanto mais amplamente do que qualquer outra prerrogativa, a máxima acima se consolidou no background filosófico por força da preponderância dos próprios modelos levados a cabo no âmbito da ciência aplicada (dir-se-ia: mais robustos e falseáveis), o que já se fazia notar pelo menos cem anos antes da popularização do monismo materialista por meio do lançamento de O erro de Descartes (DAMASIO, 1994); para um modelo inovador de tipo mente/sistema nervoso da primeira metade do século XIX, ver Bell (1974).
Curiosamente, o aprofundamento desta tendência, no âmbito dos estudos da mente, fez-se antirreducionista, propiciando uma paisagem conceitual cuja aparência mais geral (superficial) parece remontar aos cânones idealistas e à dicotomia mente-corpo: os neurônios (bem como a glia) são células estereotipadas; a transmissão é de tipo “tudo ou nada”, sendo, pois, mediada pela capacidade de armazenamento e liberação das vesículas sinápticas (KANDEL ET AL., 2000).
Partindo deste ponto – e contrariamente à afirmação da imaterialidade do cogito –, desponta a tendência de considerar a conectividade como leitmotiv da realidade operacional e fenomenológica da vida mental. Na atualidade, este paradigma se encontra amplamente endossado tanto por análises in situ quanto por simulações computacionais, o que corrobora o fato de despontar como derradeiro princípio unificador das diversas aptidões cognitivas, a despeito da elegância teórica de alguns paradigmas alternativos, como o paradigma da coativação quântica (PENROSE, 1989) e a existência pulverizada de outros menos elegantes ou mais obscuros.
Neste sentido, uma classe de projetos epistemológicos denominada Teorias Gerais da Cognição desenvolveu-se para a delimitação das propriedades essenciais dos sistemas inteligentes, tanto em termos estruturais quanto procedurais. Neste âmbito, ganhou independência uma subclasse de estudos relativos às propriedades estruturais e procedurais típicas dos organismos biológicos, a subclasse das “arquiteturas cognitivas de caráter biológico”(referidas apenas como “arquiteturas cognitivas”), a qual manteremos em foco no presente estudo (para uma introdução aos fundamentos das Teorias Unificadas da Cognição e ao conceito de arquitetura cognitiva, ver Newell (1990)).
Posto que “arquiteturas cognitivas” se referem a existências reais (ou simulações), é natural que se escore em prerrogativas evolucionárias. Em outras palavras, podemos considerar arquiteturas cognitivas como causas proximais da atividade cognitiva, as quais, por sua vez, demandam a modelagem de causas distais, servindo-lhes de escopo (MAYR, 1998; TINBERGEN, 1963). Destas causas distais, alguns prolegômenos acerca dos determinantes ônticos da vida mental humana se fazem indispensáveis. Segundo Newell (1990), os mais decisivos para a modelagem de arquiteturas cognitivas são: somos seres voltados a objetivos; dependentes de bancos de dados grandes e potencialmente não degradáveis; inexoravelmente caracterizados pelo uso de símbolos/abstrações e da flexibilidade operacional assim atingida; além de nos caracterizarmos por uma capacidade biologicamente inusual de aprender, alinhada à inusual riqueza de estímulos do ambiente adaptativo da espécie.
Delineada a concepção de Newell (1990), é preciso ter em vista que a derradeira consolidação dos prolegômenos ônticos subjacentes à arquitetura cognitiva humana permanece em discussão. Especificamente, é de se notar que, direta ou indiretamente, certos modelos privilegiam concepções teleológicas da mente, tratando como erro ou exceção os outputs passíveis de romper com os axiomas da racionalidade, seja em seu caráter operacional (ou seja, para Newell e colaboradores o comportamento é uma função de um plano objetivo prévio) ou enquanto vetores da maximização da Função Utilidade (para uma definição de axiomas racionalistas e da função utilidade, ver Von Neumann e Morgenstern (1944)).
Com estes princípios em vista, introduziremos uma breve história das arquiteturas cognitivas e, em seguida, realizaremos uma revisão sistemática e discussão crítica de um dos mais consistentes dispositivos da atualidade: o princípio dos “pequenos mundos”. Por fim, consideramos o potencial deste dispositivo para a psicopatologia, por meio de considerações endereçadas à etiologia e nosologia da esquizofrenia, o que se justifica dado seu caráter não trivial no que tange à associação de ideias (BLEULER, 1911) e desvios em relação aos padrões normais de agenciamento e interpretação da atividade mental, como é o caso para os delírios e ilusões (FRITH, 2005).
Breve História das Arquiteturas Cognitivas: Modelos Físico-simbólicos e Modelos Conexionistas
Sob o princípio da inexorabilidade mente/sistema nervoso, já na primeira metade do século XX os modelos de processamento informacional em sistemas biológicos passaram a absorver profundas influências da teorização dos processadores binários, que, tendo se iniciado no âmbito da matemática, tornaram-se popular por meio do primeiro computador digital operável, o ENIAC, e seu sucessor, o EDVAC (para uma revisão deste histórico, ver Goldstine (1972)).
Tal como se aplica aos computadores digitais, fez-se paradigmático considerar que a mente poderia ser descrita como um sistema que realiza operações finitas por força de encadeamentos potencialmente infinitos, isto é, uma máquina de Turing (para conhecer a máquina de Turing, ver Herken (1995); para uma revisão, ver Cummins (1989)). Esta tendência, por sua vez, deu origem às arquiteturas físico-simbólicas de tipo biológico (doravante referidas como “simbólicas”), cujo apogeu acompanhou de perto o desenvolvimento das diretrizes em Inteligência Artificial, sobretudo no que tange às linguagens de programação (para conhecer a mais influente arquitetura cognitiva de inspiração simbólico-computacional da história, ver Laird ET AL., 1987).
Muitos foram os acréscimos efetuados ao longo da história das arquiteturas simbólicas (para um modelo clássico, ver Newell e Simon (1972); para uma revisão, ver Johnson-Laird (1988)). Considerando as principais contribuições, podemos sumariar que a mente seria a propriedade emergente da coexistência de módulos cognitivos integrados por um processador central, o qual daria um aspecto geral serialista à arquitetura cognitiva. A formação de conteúdos mentais dar-se-ia por força de regras prévias de funcionamento, por meio das quais a experiência do “sentido” representaria uma função da ordem simbólica (semântica emergente da sintaxe).
Esta sintaxe, inacessível à introspecção (“metassintaxe”), abrigaria os componentes da vida mental de maneira fundamentalmente proposicional e enxuta, divergindo da sintaxe das línguas naturais e sendo, por isso, frequentemente denominada “linguagem do pensamento” ou “mentalês” (para conhecer um modelo clássico, ver Fodor (1997), Fodor e Pylyshyn (1988) e Shagrir (1997); para uma revisão, ver Searle (1990) e também Shagrir (1997)).
Por meio desses princípios, os estudos em arquiteturas simbólicas ganharam proeminência no âmbito da linguística e da filosofia da linguagem, a qual, de certo modo, preservam até hoje.
Não obstante, em outros domínios da ciência cognitiva, despontaram várias e duras críticas a estas arquiteturas simbólicas. Algumas destas permanecem ainda em debate, como a da imponderabilidade da emergência da semântica desde a sintaxe (SEARLE, 1990); outras se mostraram derradeiramente contundentes, como “a insolubilidade do paradoxo da degradação suave”, o qual destaca que, se a arquitetura cognitiva fosse composta de “bytes-símbolos”, a morte de alguns deles afetaria seletivamente representações mentais e rotinas cognitivas, comprometendo o caráter operacional do sistema cognitivo naquele domínio (degradação catastrófica). Tal como ilustrado pelo divertido, mas profundo, “paradoxo da avó”, a “degradação catastrófica” não ocorre: nenhuma concussão é capaz de extinguir seletivamente a existência mental de sua avó ou a compreensão e uso do conceito de “avó” – ainda que o princípio não seja de todo alheio ao que ocorre nas agnosias, associadas ao comprometimento de populações neurais relativamente localizadas, frequentemente no córtex parietal (para uma revisão, ver Gross (2005)).
A partir destas prerrogativas e, sobretudo, da consolidação da parceria entre as neurociências e a ciência computacional ao longo da década de 1970 e início da década de 1980, modelagens do funcionamento da mente diretamente inspiradas no comportamento neural observado in situ começaram a ganhar força, em consonância ao estreitamento dos limites científico-filosóficos da ciência cognitiva, cada vez mais associada a princípios popperianos. Já na década de 1980, fazia-se notar o predomínio das modelagens conexionistas sobre as simbólicas (para uma revisão, ver Mira (2008)).
No que tange ao perfil neurofisiológico da mente, os grandes diferenciais conexionistas foram: rompimento metodológico com a relação neurônios-símbolos e com os determinantes sintáticos associados, em prol de fundamentos subsimbólicos (FRIXIONE; SPINELLI, 1992); abandono do design definido pela relação modularidade dos módulos cognitivos/centralidade do processador de códigos; paralelismo maciço; e degradação suave (SMOLENSKY, 1988). Um aspecto menos determinante, mas popularmente conhecido, é a possibilidade de representações gráficas econômicas e sistemáticas. No que tange à Teoria do Conhecimento, um aspecto de destaque é representado pelo abandono de representações localizáveis para o conhecimento, que desde então foi assumido como o resultado da operação total do sistema.
Em geral, as arquiteturas tipicamente conexionistas (denominadas “elementares”) exibem três camadas de nódulos/neurônios, em relação às quais a ativação se propaga linearmente (feed-forward): as camadas de entradas, as camadas de saídas e as camadas internas dotadas de aspecto de rede, contando com número variável de nódulos/neurônios (para uma introdução, ver Marcus (2001)). Ao longo destas redes, assume-se que o grau de ativação (tecnicamente denominado de “peso”) das conexões seja variável, conferindo sensibilidade à interface com o ambiente.
Assim, a experiência em constante processo de acumulação correlacionar-se-ia à dinâmica distributiva dos pesos conectivos, genericamente denominada “aprendizado” e intrinsecamente dependente de retroalimentações capazes de garantir um balanceamento interno apropriado. Em relação ao programa constitucional implicado nas arquiteturas simbólicas, os conceitos de condicionamento e treinamento adquirem maior importância epistemológica e o conexionismo enseja uma moderada aproximação às modalidades menos “eliminatórias” do behaviorismo (SCHRETER, 1990).
Isto é, direta ou indiretamente, assume-se que a capacidade intelectual humana seja fundamentalmente caracterizada pela plasticidade típica de nossas células nervosas em associação com a exposição suficiente do organismo aos estímulos apropriados, em face da qual o caráter de Teoria Geral da Cognição do conexionismo emerge da associação entre número de nódulos e de camadas intermediárias, pesos iniciais e dinâmica de alterações dos pesos conectivos (para uma revisão, ver Marcus (2001) e Smolensky (1990)).
Ainda que o conexionismo elementar permaneça muito influente, ao longo das últimas das décadas as limitações das arquiteturas dotadas dessas características tornaram-se claras em dois níveis. No primeiro nível, mesmo em suas formas mais bem acabadas, as arquiteturas conexionistas não se mostram muito apropriadas para a modelagem da aquisição da linguagem, da recursividade e do aprendizado por exposição única (para uma revisão destas arquiteturas “mais bem acabadas”, ver Tryon (2002); para uma revisão dessas fraquezas, ver Marcus (2001)). No segundo nível, arquiteturas inspiradas no funcionamento do sistema nervoso precisam contemplar o fato de este abarcar modularidade, por meio da implementação de áreas unimodais, transmodais e multimodais, neurônios menos e mais conectados, ligações excitatórias e inibitórias e diversos mecanismos de plasticidade seletiva (para uma introdução à modularidade e aspectos relacionados, ver Kandel (2000)).
Neste sentido, o maior conhecimento acerca da organização modular do cérebro humano, em parte propiciado pela recente evolução dos métodos de registro (isto é, FMRI, QEEG de Alta Resolução, MEG, DTI), possibilitou o estabelecimento de novas prioridades e linhas de pesquisa, tanto mais significativas para a ontologia da condição mente/sistema nervoso do que as que se faziam possíveis por meio de modelos puramente simbólicos ou conexionistas elementares do passado.
Por exemplo, como modelar arquiteturas cognitivas a partir de registros da atividade cognitiva humana? Como é que deve ser a arquitetura de um modelo da mente, para que exiba simultaneamente a propriedade de realizar tarefas específicas (isto é, identificação visual de um objeto) e se integrar em tarefas complexas (como avaliar esteticamente um objeto visto), de maneira igualmente eficiente/funcional? Quais seriam os vetores presentes nas arquiteturas cognitivas responsáveis pelas alterações anatômicas e fisiológicas do cérebro de tipo psicopatológico?
Estas são algumas das questões que se levantam em relação ao tema e que devem ser mantidas em foco quando refletimos sobre o mesmo, ainda que na ocasião não as possamos esgotar. Ainda que atualmente estas “prioridades” permaneçam mais associadas a novas perguntas do que a novas respostas, uma prerrogativa já desponta com promissora exuberância: ao menos em parte, a mente humana obedece a princípios de “pequenos mundos” (small worldness) e às propriedades conectivas entre estes. É deste tópico que trataremos a seguir.
Galáxia de Pequenos Mundos
Nas seções a seguir, iremos considerar uma tarefa tripla: na seção 2 destrincharemos o princípio de “pequenos mundos”, esclarecendo seu papel para o presente e o futuro das modelagens mente/sistema nervoso; na seção 3, consideraremos uma revisão sistemática da organização do campo; na seção 4, o foco será o potencial do princípio de pequenos mundos para o aprofundamento de análises psicopatológicas, especialmente no que tange à etiologia de desordens associadas a marcadores endofenotípicos, para o que tomaremos a esquizofrenia como mote.
Introdução ao Conceito de “Pequenos Mundos” (Small Worldness) e seu Papel para o Entendimento da Mente
O conceito de pequenos mundos não é novo, descrevendo de maneira geral a organização de redes com múltiplos nódulos, em que a distância entre quaisquer dois seja curta, de modo a atender simultaneamente a demanda por otimização local e global (YU ET AL., 2008). Sua aplicação pode servir a problemáticas tão diversas quanto a modelagem da relação entre os sites da web (WWW) e experimentos de psicologia social.
A alcunha “pequenos mundos” foi cunhada no famoso experimento dos “seis graus de separação”, em que sujeitos eram requisitados a enviar pacotes pelo correio, os quais deveriam chegar a remetentes que eles desconheciam, por meio de elos intermediários, que se revelaram menores do que seis, em todas as ocasiões em que o pacote chegou ao seu destino final (MILGRAM, 1967).
No âmbito da elucidação da condição mente/sistema nervoso, o conceito frequentemente se associa aos de distribuição funcional dos neurônios e conexões e propriedades não escalares das networks biológicas. Estes, por sua vez, retroagem ao fato de estas networks poderem ser descritas tal como se obedecessem a parâmetros escalares denominados “leis de força” (power laws), que descreveremos a seguir e que se associam à caracterização das relações conectivas, a meio termo entre o estabelecimento de ligações locais funcionais e a funcionalidade das ligações longas, características da integração modular das subpopulações neurais.
A primeira aplicação do conceito para o desenvolvimento de arquiteturas cognitivas data de pouco mais de dez anos (WATTS; STROGATZ, 1998) e ainda hoje permanece a mais famosa e citada. Não obstante, o que esta aplicação representa para o entendimento da mente depende de uma etapa de definição de conceitos elementares, em relação à qual nos pautaremos pelas lições de Barabási e Oltvai (2004); posto que estas lições versam sobre arquiteturas biológicas em geral, nós tomaremos a liberdade de adaptá-las ao presente foco de interesse.
2.a. Denomina-se k o grau de conectividade de um nódulo/neurônio, sendo 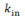 o número de entradas conectivas e
o número de entradas conectivas e 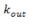 o número de saídas conectivas. Dada uma network cujos nódulos e conexões sejam distribuídos a esmo, muitos nódulos terão o mesmo k que a média da network. Porém, em sistemas biológicos complexos como a mente humana, isso não ocorre, sendo que a distribuição de k segue a regra P(k) ~
o número de saídas conectivas. Dada uma network cujos nódulos e conexões sejam distribuídos a esmo, muitos nódulos terão o mesmo k que a média da network. Porém, em sistemas biológicos complexos como a mente humana, isso não ocorre, sendo que a distribuição de k segue a regra P(k) ~ 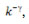 , em que P(K) expressa a probabilidade de um determinado nódulo/neurônio possuir k conexões
, em que P(K) expressa a probabilidade de um determinado nódulo/neurônio possuir k conexões 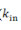 +
+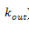 , ~ significa “proporcional a” e
, ~ significa “proporcional a” e 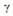 representa o grau do expoente. Esta power law expressa que, quanto menor o valor de
representa o grau do expoente. Esta power law expressa que, quanto menor o valor de 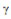 , mais importantes se tornam os neurônios mais conectados e, quanto maior
, mais importantes se tornam os neurônios mais conectados e, quanto maior 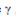 , menos hierarquia apresentará o sistema (BARABÁSI; OLTVAI, 2004).
, menos hierarquia apresentará o sistema (BARABÁSI; OLTVAI, 2004).
2.b. Para os sistemas biológicos complexos, 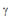 entre 2 e 3 (BARABÁSI; ALBERT, 1999), este valor expressa matematicamente o fato de que os nódulos nem possuem distribuição conectiva passível de ser reduzida a modelos de distribuição randômica (em que o k total se faz a medida comum), nem organizações hierárquicas estritamente centradas no papel de alguns neurônios extremamente conectados (BARABÁSI; OLTVAI, 2004).
entre 2 e 3 (BARABÁSI; ALBERT, 1999), este valor expressa matematicamente o fato de que os nódulos nem possuem distribuição conectiva passível de ser reduzida a modelos de distribuição randômica (em que o k total se faz a medida comum), nem organizações hierárquicas estritamente centradas no papel de alguns neurônios extremamente conectados (BARABÁSI; OLTVAI, 2004).
2.c. A arquitetura da condição mente/sistema nervoso implica a existência de alguns neurônios hiperconectados (hubs), primariamente ligados a células tanto menos conectadas e secundariamente a outros hubs.
2.d. Padrões gerais de conectividade podem ser denominados “motivos” (motifs). Dado o alto de grau de conservação presente nos designs funcionais (isto é, dado o caráter conservador da evolução), os motivos repetem-se como padrões ao longo do cérebro. Eles são característicos da modularidade, cuja implementação se dá por meio de padrões francamente recorrentes.
Agora passaremos às considerações mais centradas nas propriedades específicas da mente, para o que nos inspiraremos no modelo de Watts e Strogatz (1998).
2.e. Considerando uma rede de nódulos cuja função seja representar a conectividade mielinizada do cérebro todo (substância branca), denominemos n o número de vértices e k o número de arestas por vértice; conectando-se aleatoriamente os vértices com probabilidade p, forma-se uma rede de conectividade aleatória (p = 1); conectando-se regularmente os vértices (qualquer que seja o algoritmo usado para tanto), forma-se uma rede cujo grau de aleatoriedade é nulo (p = 0). Dadas estas definições, a arquitetura da mente/sistema nervoso obedece a padrões de conectividade em que 1 > p > 0.
2.f. Denominado L(p) o comprimento dos axônios individuais e denominado C(p) o coeficiente de aglutinação dos inputs sinápticos dos nódulos/neurônios das camadas intermediárias e de saída, de modo que a distância absoluta entre todos os axônios da rede representante da conectividade subjacente ao fenômeno mente/sistema nervoso seja determinada pelo L(p) total, e o grau de agrupamento total dos nódulos/neurônios, pelo C(p) total, revela-se a existência de grandes intervalos de p nos quais L(p) é quase-randômico, ao passo que o coeficiente de aglutinação C(p) permanece maior do que randômico. Isso sugere a existência de áreas de concentração conectadas por “atalhos” a outras áreas de concentração, dando à arquitetura cognitiva características topológicas de pequenos mundos organizados em “motivos”.
2.g. Não são necessários muitos atalhos conectivos de longa distância para dar a uma rede conectiva de tipo mente/sistema nervoso características de pequenos mundos.
Agora passaremos para considerações gerais:
2.h. Os neurônios hiperconectados (hubs), caracterizados por valores de k altos, estão principalmente dispostos nas áreas corticais de associação multimodal. Especificamente, os maiores valores de k estão em populações neurais das áreas de Broadman BA 6;11;21;29;46;14 (BASSETT ET AL., 2008).
2.i. Os hubs existem em pequenas quantidades (YU ET AL., 2008).
2.j. No cérebro humano, as áreas de associação multimodais possuem as conexões mais longas (altos valores de L(p)), seguidas pelas unimodais e então pelas áreas transmodais, destacando-se o fato de que este é um achado aparentemente intrigante, explicável na medida em que tenhamos em mente que muitas das áreas transmodais (isto é, sistema límbico) são filogeneticamente antigas e que as áreas unimodais estão associadas por conexões multimodais. Funções executivas caracterizam-se por k alto e C baixo (BASSETT ET AL., 2008).
2.k. Por meio dos atalhos conectivos às áreas corticais de associação multimodal, o alto grau de aglutinação em sítios específicos resolve-se em atividade ordenada, de modo tal que a arquitetura conectiva humana se assemelha a uma rede de pequenas redes. Ao contrário de arquiteturas conexionistas elementares, com sua convergência global à conformação uniforme, a conectividade da mente humana mais se aproxima estruturalmente a uma galáxia, mantida coesa por meio das relações atrativas e repulsivas dos “pequenos mundos”, ou à WEB (www), na qual proliferam sites pequenos, preferencialmente com links para sites maiores (BARABÁSI; OLTVAI, 2004).
2.l. É possível conceber arquiteturas cognitivas à luz de diferenciações conjunturais entre “tijolos” (bricks), isto é, as células implicadas na composição dos motivos, e “pontes” (bridges), isto é, as vias conectivas principais. Não obstante, estes aspectos nem sempre podem ser distinguidos com precisão (HUANG ET AL., 2007).
2.m. O disparo ordenado e frequentemente sincrônico de potenciais de ação em áreas distintas do sistema nervoso humano, integradas pela execução de uma função ou tarefa e assim produzindo coerência em faixas diversas do EEG (retina, núcleo geniculado do tálamo, córtex occipital primário, córtex ocipital secundário), obedece a princípios arquitetônicos de pequenos mundos (YU ET AL., 2008). Em outras palavras, redes com propriedades de pequenos mundos são tão dinâmicas quanto as aptidões dinâmicas do sistema.
Figura 1
Modelo computacional de networks cognitivas em primatas, cuja arquitetura é inspirada no princípio de pequenos mundos
Fonte: Stephan et al. (2000)
Revisão Sistemática dos Achados Recentes
As palavras-chave small world network, small world and cognitive architecture, small world and brain e small worldness resgatam milhares de resumos de artigos publicados entre 2007 e 2009, no Pubmed (<http://www.ncbi.nlm.nih.gov/pubmed>) e no ISI (<http://apps.isiknowledge.com>).
O resgate dos artigos integrais (graças ao convênio: <http://www.usp.br/sibi/>) revela 87 artigos abordando diretamente arquiteturas de pequenos mundos relacionadas ao cérebro humano.
Uma nova seleção, considerando apenas manuscritos que 1) tratem de arquiteturas corticais (excluindo estudos em populações neurais específicas), 2) tratem de cognição (exclusão de sistemas sensoriais e emoções) e 3) tenham sido publicados em revistas indexadas no ISI, na área de psicologia/neurociências (excluindo revistas de física, matemática e computação), revela que 37 publicações (referências em anexo) são responsáveis pela organização do campo em oito tópicos de interesse:
•small world e EEG;
•condensação nodular e distância conectiva;
•funcionalidade conectiva do cérebro humano;
•sincronia populacional de neurônios em redes de pequeno mundo;
•robustez das redes neurais explicada por princípios de pequenos mundos;
•genética das redes de pequenos mundos;
•métodos quantitativos para a determinação de caráter de pequeno mundo de arquitetura cognitiva; e
•FMRI e pequenos mundos.
Figura 2
Organização do Campo de Estudos “Cognição e Pequenos Mundos”*

Obs.: * Este campo se caracteriza por uma relativa heterogeneidade dos estudos. Os pesquisadores formam grupos pequenos de pesquisa, que colaboram pouco entre si em tópicos de interesse comum.
Esta distribuição sugere que:
3.a. A perspectiva fundamental do campo é a aglutinação conectiva e a distribuição das células. Alguns destes estudos se relacionam aos de migração neuronal e regulação epigênica. Regulação genética das arquiteturas cognitivas é um assunto pouco explorado.
3.b. A ausência de degradação catastrófica no âmbito da arquitetura cognitiva humana vem sendo associada às propriedades de pequenos mundos.
3.c. Estudos de imageamento estão baseados em FMRI; isto significa que o campo ainda carece de estudos endereçando a substância branca diretamente (DTI e, recentemente, q-Ball) para difusão de tensor em múltiplas fibras.
3.d. Número considerável de estudos aborda momentos de sincronia e dessincronia eletroencefalográfica (epochs) como reflexo da arquitetura de pequenos mundos e alterações em seus parâmetros momentâneos.
3.e. Considerações desenvolvimentais acerca da consolidação do endofenótipo das arquiteturas cognitivas maduras ainda não foram abordadas.
O Conceito de Pequenos Mundos Aplicado à Compreensão da Etiologia e Prognóstico de Desordens Mentais com Componentes Endofenotípicos
Atualmente, despontam evidências de que diversas desordens mentais (esquizofrenia, Alzheimer, epilepsia, doença de Parkinson) expressam distúrbios em redes de pequeno mundo (STAM; REIJNEVELD, 2007). Nesta seção, iremos apresentar os resultados de uma revisão dos estudos em “esquizofrenia e alterações topológicas de pequenos mundos”, conduzida por meio da aplicação dos mesmos filtros definidos na terceira seção para o período 2006-2009, em relação à qual consideraremos privilegiadamente o modo como estas novas perspectivas influenciam modelos etiológicos e nosológicos.
Arquitetura Cognitiva na Esquizofrenia
A esquizofrenia é uma síndrome e não exatamente uma doença, na medida em que seus sintomas não são totalmente definidos. Ainda assim, despontam evidências estatísticas da relação entre diagnóstico de esquizofrenia e déficits cognitivos, eventualmente relacionados por meio de alterações morfofuncionais e anatomofuncionais do cérebro (LAWRIE ET AL., 2008).
Entre estas alterações, um dos principais fatores de risco são as alterações na substância branca (ASHTARI ET AL., 2007; BUCHSBAUM ET AL., 1998; BURNS ET AL., 2003; DAVIS ET AL., 2003; DEARY ET AL., 2006), recentemente ligadas à exocitose glutamatérgica, que é neurotóxica e exibe potencial para afetar a mielinização axônica (ZISKIN ET AL., 2007). Também recentemente, despontaram estudos associando tal achado funcional a polimorfismos do gene Neurogelin-1, o qual participa das cascatas bioquímicas que convergem para a síntese dos receptores glutamatérgicos de tipo NMDA e é um dos marcadores genéticos mais consistentes da síndrome (para uma revisão destes achados, ver HALL ET AL. (2006), KONRAD; WINTERER (2008) e ROY ET AL. (2007)).
Dentro deste panorama, é possível que existam relações entre o solipsismo de cunho esquizofrênico (caracterizado por interpretações idiossincráticas de discursos e ações, retração social, negativismo, entre outros) e desconexões executivas, por sua vez responsáveis por uma perda geral na capacidade de agir intencionalmente (KAISER; WEISBROD, 2007).
Paralelamente, destacam-se evidências de que alterações da substância branca se relacionem à manifestação de alucinações auditivas. Uma possibilidade é que isso ocorra por meio da supressão do mecanismo responsável pela capacidade de diferenciar subvocalizações de inputs auditivos, pela transmissão de cópias eferentes dos pensamentos verbalizados (subvocalizações) para as áreas sensoriais do córtex temporal, dessensibilizando-o. Tal como o ato motor dessensibiliza o agente intencional que, por exemplo, é incapaz de provocar cócegas em si mesmo, este mecanismo (corollary discharge) daria substrato para a identificação da diferença fenomenológica entre estímulos verbais externos e internos, a qual se perderia junto a alterações funcionais na transmissão frontotemporal (FRISTON; FRITH, 1995; FRITH, 2005).
Em relação a essas hipóteses, podemos alinhavar estudos que sugerem que alterações no valor médio do coeficiente de aglutinação conectiva (C), bem como alterações nos padrões estatisticamente normais de distribuição dos hubs, isto é, das célulascaracterizadas por altos valores de f, estejam relacionadas à síndrome.
Em um estudo de fRMI, no qual os coeficientes foram calculados tendo por base a topologia e o volume da substância cinzenta, Bassett et al. (2008) verificaram que:
4.a. Alguns dos principais parâmetros topológicos estão preservados nas arquiteturas cognitivas dos portadores de esquizofrenia.
4.b. Áreas transmodais e unimodais apresentam parâmetros de conectividade e relações hierárquicas correspondentes a controles.
4.c. Áreas multimodais apresentam menor conectividade hierárquica e distâncias conectivas L(p) elevadas.
Agora passaremos a considerações mais gerais:
4.d. Entre portadores de esquizofrenia, os hubs tendem a revelar altos graus de aglutinação e a ser particularmente conectados entre si. É possível que o grau de alteração deste parâmetro esteja positivamente correlacionado à cronicidade (LIU ET AL., 2008).
4.e. Hubs tipicamente pré-frontais (em controles) tendem a ser topologicamente substituídos por hubs: temporal-inferiores, insulares e cingulares.
Entrementes, permanece ainda incerto como e em que grau os valores paramétricos das arquiteturas cognitivas de pequenos mundos dos portadores de esquizofrenia se diferenciam daqueles encontrados em controles. Apenas dois estudos abordaram este tópico diretamente: de acordo com Bassett et al. (2008), os valores são equivalentes; de acordo com Liu et al. (2008), eles são diferentes.
4.f. A velocidade média de transmissão de informações entre regiões diversas do sistema nervoso central encontra-se diminuída na esquizofrenia (LIU ET AL., 2008).
4.g. A coerência intra e inter-hemisférica nas bandas alfa, beta e gama apresenta-se diminuída no EEG de portadores de esquizofrenia (MICHELOYANNIS ET AL., 2006), havendo sugestões de que esta perda de funcionalidade das arquiteturas cognitivas córtico-corticais se sobressaia mais pela execução de tarefas cognitivas simples do que pela execução de tarefas complexas (PACHOU ET AL., 2008).
4.h. Em situações de descanso (olhando para um ponto na parede), a topologia da arquitetura cognitiva dos portadores de esquizofrenia, obtida por meio de EEG, sugere um aumento no número de padrões de atividade sincrônica mediados por conexões randômicas, o que pode se relacionar às particularidades sintomatológicas dos portadores da síndrome (RUBINOV ET AL., 2009).
A associação destes aspectos sugere que a esquizofrenia se caracterize por perdas na conformação dos padrões de motivos e conectividade hierárquica, dependentes de graus consideráveis de controle inibitório e excitatório de origem pré-frontal. Mormente, destacam-se eventuais diminuições nas taxas de transferências de informações, isto é, há alterações de sinalização neural e perdas de sincronia em bandas relacionadas a tarefas executivas (com destaque para Beta 1 e Beta 2), possivelmente implicadas em sintomas cognitivos, e variações de QI em relação a controles normais.
Por fim, considerando o fato de que diminuições no volume total da substância cinzenta representam um dos achados mais consistentes no que tange à patofísica da esquizofrenia (PANTELIS ET AL., 2005), é de se supor que um dos fatores relacionados aos achados topológicos seja a perda de neurônios em determinadas áreas críticas do córtex em momentos críticos do desenvolvimento (eventualmente por poda neural excessiva), a partir dos quais a conectividade sofra conformações crescentemente diversas daquelas encontradas em controles.
Ainda não existem estudos abordando esta conjuntura, até porque ainda se fazem necessários modelos consistentes (derivados de estudos longitudinais) relativos às alterações topológicas das arquiteturas cognitivas ao longo do desenvolvimento em sujeitos normais, para que então se determinem as relações entre a quantificação das alterações anatômicas e variações esperadas nas arquiteturas cognitivas em portadores de esquizofrenia e outras populações. Não obstante, é possível se aventar que achados desta natureza sucedam os presentes.
CONSIDERAÇÕES FINAIS
Ao longo de sua curta história, as arquiteturas cognitivas de tipo mente/sistema nervoso evoluíram em sentido ao conhecimento anatomofisiológico acumulado acerca da estrutura do sistema nervoso humano e, recentemente, em sentido à psicopatologia.
É muito provável que esta tendência se torne cada vez mais forte, de modo que se faz interessante aos psicólogos e cientistas afins aprofundar seus conhecimentos neste domínio. Enfim, após esta revisão, conclui-se que a modelagem do modo de processamento informacional do cérebro humano emerge como um projeto viável, baseado em modelos computacionais que finalmente parecem se adequar à verdadeira complexidade de nossa arquitetura cognitiva. Este panorama sugere um papel fundamental para a psicologia, o qual, diga-se de passagem, alinha-se à possibilidade de tratamento das desordens nervosas por meio da introdução de precursores ou células, em áreas específicas do cérebro, e às novas possibilidades de prognóstico que devem despontar do tratamento de transtornos do sistema nervoso central, os quais devem introduzir potenciais desenvolvimentos associados a um sem-número de novos desafios.
REFERÊNCIAS
ASHTARI, M.;COTTONE, J.; ARDEKANI, B. A.; CERVELLIONE, K.; SZESZKO, P. R.; WU, J.; STEVEN, C.; SANJIV, K. Disruption of white matter integrity in the inferior longitudinal fasciculus in adolescents with schizophrenia as revealed by fiber tractography. Archives of General Psychiatry, v. 64, n. 11, p. 1.270-1.280, 2007. [ Links ]
BARABÁSI, A. L.; ALBERT, R. Emergence of scaling in random networks. Science, n. 289, p. 509-512, 1999. [ Links ]
______; OLTVAI, Z. N. Network biology: Understanding the cell’s functional organization. Nature Reviews Genetics, v. 5, n. 2, p. 101-113, 2004.
BASSETT, D. S.;BULLMORE, E.; VERCHINSKI, B. A.; MATTAY, V. S.; WEINBERGER, D. R.; MEYER-LINDENBERG, A. Hierarchical organization of human cortical networks in health and schizophrenia. The Journal of Neuroscience, v. 28, n. 37, p. 239-248, 2008. [ Links ]
BELL, C. Idea of a new anatomy of the brain. In: CRANEFIELD, P. (Ed.). Francois Magendie, Charles Bell and the Course of the Spinal Nerves. New York: Futura, 1974.
BLEULER, E. Dementia praecox or the group of schizophrenias. New York: International University Press, 1911. [ Links ]
BUCHSBAUM, M. S; TANG, C. Y.; PELED, S.; GUDBJARTSSON, H.; LU, D.; HAZLETT, E. A.; DOWNHILL, J.; HAZNEDAR, M.; FALLON, J. H.; ATLAS, S. W. MRI white matter diffusion anisotropy and PET metabolic rate in schizophrenia. NeuroReport, v. 9, n. 3, p. 425-430, 1998. [ Links ]
BURNS, J.; JOB, D.; BASTIN, M. E.; WHALLEY H.; MACGILLIVRAY, T.; JOHNSTONE, E. C.; LAWRIE, S. M. Structural disconnectivity in schizophrenia: a diffusion tensor magnetic resonance imaging study. The British Journal of Psychiatry, v. 182, n. 5, p. 439-443, 2003. [ Links ]
CHURCHLAND, P. S. Neurophilosophy: toward a unified science of the mind/brain. Cambridge: MIT Press, 1986. [ Links ]
CUMMINS, R. Meaning and mental representation. Cambridge: The MIT Press, 1989. [ Links ]
DAMASIO, A. Descartes’ error: emotion, reason, and the human brain. New York: Avon, 1994.
DAVIS, K. L.; STEWART, D. G,; FRIEDMAN, J. I.; BUCHSBAUM, M.; HARVEY, P. D.; HOF, P.R,; BUXBAUM, J.; HAROUTUNIAN, V. White matter changes in schizophrenia: evidence for myelin-related dysfunction. Archives of General Psychiatry, v. 60, n. 5, p. 443-456, 2003. [ Links ]
DAVIS, M. The undecidable. Basic papers on undecidable propositions, unsolvable problems, and computable functions. New York: Raven Press, 1965. [ Links ]
DEARY, I. J; BASTIN, M. E.; PATTIE, A.; CLAYDEN, J. D.; WHALLEY, L. J.; STARR, J. M.; WARDLAW, J. M. White matter integrity and cognition in childhood and old age. Neurology, v. 66, n. 4, p. 505-12, 2006. [ Links ]
EDELMAN, G. Neural darwinism: selection and reentrant signaling in higher brain function. Neuron, v. 10, n. 2, p. 115-125, 1993. [ Links ]
FODOR, J. Connectionism and the problem of systematicity (continued): why Smolensky’s solution still doesn’t work. Cognition, v. 62, n. 1, p. 109-119, 1997.
______; PYLYSHYN, Z. W. Connectionism and cognitive architecture: a critical analysis. Cognition, v. 28, n. 1-2, p. 3-71, 1988. [ Links ]
FRISTON, K. J.; FRITH, C. D. Schizophrenia: a disconnection syndrome? Clinical neuroscience, New York, v. 3, n. 2, p. 89-97, 1995. [ Links ]
FRITH, C. The neural basis of hallucinations and delusions. Comptes Rendus Biologies, v. 328, n. 2, p. 169-175, 2005. [ Links ]
FRIXIONE, M.; SPINELLI, G. Connectionism and functionalism: the importance of being a subsymbolist. Journal of Experimental & Theoretical Artificial Intelligence, v. 4, n. 1, p. 3-17, 1992. [ Links ]
GOLDSTINE, H. H. The computer from Pascal to von Neumann. Princeton: Princeton University Press, 1972. [ Links ]
GROSS, C. G. Genealogy of the “Grandmother Cell”. The Neuroscientist, v. 8, n. 5, p. 512-518, 2005.
HALL, J.; WHALLEY; H. C.; HEATHER C.; JOB, D. E.; BAIG, B. J.; MCINTOSH, A. M.; EVANS, K. L.; THOMSON, P. A.; PORTEOUS, D. J.; CUNNINGHAM-OWENS, D. G.; JOHNSTONE, E. C.; LAWRIE, S. M. A neuregulin 1 variant associated with abnormal cortical function and psychotic symptoms. Nature Neuroscience, v. 9, n. 12, p. 1.477-1.478, 2006. [ Links ]
HERKEN, R. (Ed.). The universal turing machine: a half-century survey. New York: Springered. 1995. [ Links ]
HUANG, C.-Y; CHENG, C-Y; SUN, C-T. Bridge and brick network motifs: identifying significant building blocks from complex biological systems. Artificial Intelligence in Medicine, v. 41, n. 2, p. 117-127, 2007. [ Links ]
JOHNSON-LAIRD, P. N. The computer and the mind. Cambridge: Harvard University Press, 1988. [ Links ]
KAISER, S.; WEISBROD, M. Intentionality as a link between the neuropsychology and the symptoms of schizophrenia. Psychopathology, v. 40, n. 4, p. 221-228, 2007. [ Links ]
KANDEL, E.;SCHWARTZ, J. H.; JESSELL, T. M. Principles of neuroscience. New York: McGraw-Hill, 2000. [ Links ]
KONRAD, A.; WINTERER, G. Disturbed structural connectivity in schizophrenia primary factor in pathology or epiphenomenon? Schizophrenia Bulletin, v. 34, n. 1, p. 72-92, 2008. [ Links ]
LAIRD, J. E.; NEWELL, A.; ROSENBLOOM P. S.Soar: An architecture for general intelligence. Artificial Intelligence, n. 33, p. 1-64, 1987. [ Links ]
LAWRIE, S. M.; MCINTOSH, A. M.; HALL, J.; OWENS, D. G.C.; JOHNSTONE, E. C. Brain structure and function changes during the development of schizophrenia: the evidence from studies of subjects at increased genetic risk. Schizophrenia Bulletin, v. 34, n. 2, p. 330-340, 2008. [ Links ]
LIU, Y.; LIANG, M.; ZHOU, Y.; HE, Y.; HAO, Y.; SONG, M.; YU, C.; LIU, H.; LIU, Z.; JIANG, T. Disrupted small-world networks in schizophrenia. Brain, v. 131, n., p. 945-961, 2008. [ Links ]
MARCUS, G. F. The algebraic mind: integrating connectionism and cognitive science. Cambridge: MIT Press, 2001. [ Links ]
MAYR, E. O desenvolvimento do pensamento biológico. Brasília: Ed. UnB, 1998. [ Links ]
MICHELOYANNIS, S.; PACHOU, E.; STAM, C. J.; BREAKSPEAR, M.; BITSIOS, P.; VOURKAS, M.; ERIMAKI, S.; ZERVASKIS, M. Small-world networks and disturbed functional connectivity in schizophrenia. Schizophrenia Research, v. 87, n. 1-3, p. 60-66, 2006. [ Links ]
MILGRAM, S. The small-world problem. Psychology Today, n. 2, p. 61-67, 1967. [ Links ]
MIRA, J. M. Symbols versus connections: 50 years of artificial intelligence. Neurocomputing, v. 71, n. 4-6, p. 671-680, 2008. [ Links ]
NEWELL, A. Unified theories of cognition. Cambridge: Harvard University Press, 1990. [ Links ]
______; SIMON, H. A. Human problem solving. Englewood Cliffs: Prentice Hall, 1972. [ Links ]
PACHOU, E.; VOURKAS, M.; SIMOS, P.; STAM, C. J.; TSIRKA, V.; MICHELOYANNIS, S. Working memory in schizophrenia: An EEG study using power spectrum and coherence analysis to estimate cortical activation and network behavior. Brain Topography, 2008. [ Links ]
PANTELIS, C.; YÜCEL, M.; WOOD, S. J.; VELAKOULIS, D.; SUN, D.; BERGER, G.; STUART, G. W.; YUNG, A.; PHILLIPS, L.; MCGORRY, P. D. Structural brain imaging evidence for multiple pathological processes at different stages of brain development in schizophrenia. Schizophrenia Bulletin, v. 31, n. 3, p. 672-696, 2005. [ Links ]
PENROSE, R. The emperor’s new mind: concerning computers, minds, and the laws of physics.Oxford: Oxford University Press, 1989.
RUBINOV, M.; KNOCK, S. A.; STAM, C. J.; MICHELOYANNIS, S.; HARRIS, A. W. F.; WILLIAMS, L. M.; BREAKSPEAR, M. Small-world properties of nonlinear brain activity in schizophrenia. Human Brain Mapping, v. 30, n. 2, p. 403-416, 2009. [ Links ]
ROY, K.; MURTIE, J. C.; EL-KHODOR, B.F.; EDGAR, N.; SARDI, S. P.; HOOKS, B. M.; BENOIT-MARAND, M.; CHEN, C.; MOORE, H.; O'DONNELL, P.; BRUNNER, D.; CORFAS, G. Loss of erbB signaling in oligodendrocytes alters myelin and dopaminergic function, a potential mechanism for neuropsychiatric disorders. Proceedings of the National Academy of Sciences, v. 104, n. 19, p. 8.131-8.136, 2007. [ Links ]
SCHRETER, Z. Connectionism-a link between psychology and neuroscience? In: ______. Symbols Versus Neurons. IEE Colloquium p. 3-7, 1990. [ Links ]
SEARLE, J. R. Is the brain a digital computer? Proceedings and Addresses of the American Philosophical Association, v. 64, n. 3, p. 21-37, 1990. [ Links ]
SHAGRIR, O. Two dogmas of computationalism. Minds and Machines, v. 7, p. 321-344, 1997. [ Links ]
SMOLENSKY, P. On the proper treatment of connectionism. Behavioral and Brain Sciences, v. 11, p. 1-74, 1988. [ Links ]
______. Tensor product variable binding and the representation of symbolic structures in connectionist systems. Artificial Intelligence, v. 46, n. 1-2, p. 159-216, 1990. [ Links ]
STAM, C. J.; REIJNEVELD, J. C. Graph theoretical analysis of complex networks in the brain. Nonlinear Biomedical Physics, v. 1, n. 1, p. 3, 2007. [ Links ]
STEPHAN, K. E.; HILGETAG C. C.; BURNS G. A.; O'NEILL M. A.; YOUNG M. P.; KOTTER R. Computational analysis of functional connectivity between areas of primate cerebral cortex. Philosophical Transactions of the Royal Society B: Biological Sciences, v. 355, p. 111–126, 2000.
TINBERGEN, N. On aims and methods of ethology. Oxford: Zeitschrift für Tierpsychologie, 1963. [ Links ]
TRYON, W. W. Contributions of connectionism to postmodern psychology. American Psychologist, v. 57, n. 6-7, p. 455-456, 2002. [ Links ]
VON NEUMANN, J.; MORGENSTERN, O. Theory of games and economic behavior. Princeton: Princeton University Press, 1944. [ Links ]
WATTS, D. J.; STROGATZ, S. H. Collective dynamics of “small-world” networks. Nature, v. 393, n. 6.684, p. 440-442, 1998.
YU, S.; HUANG, D.; SINGER, W.; NIKOLIC, D.A small world of neuronal synchrony. Cerebral Cortex, v. 18, n. 12, p. 2.891-2.901, 2008. [ Links ]
ZISKIN, J. L.; NISHIYAMA, A.; RUBIO, M.; FUKAYA, M.; BERGLES, D. E. Vesicular release of glutamate from unmyelinated axons in white matter. Nature Neuroscience, v. 10, n. 3, p. 321-330, 2007. [ Links ]
 Endereço para correspondência
Endereço para correspondência
Álvaro Machado Dias
E-mail:alvaromd@usp.br
Submetido em: 04/09/2009
Revisto em: 08/02/2010
Aceito em: 27/02/2010