










Serviços Personalizados
Journal

artigo
 Português (pdf)
Português (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
Indicadores
Compartilhar

 Permalink
PermalinkInteramerican Journal of Psychology
versão impressa ISSN 0034-9690
Interam. j. psychol. v.40 n.2 Porto Alegre ago. 2006
ARTÍCULOS
Cooperación en ratas: efectos de la experiencia temprana
Cooperative behavior in rats: effects of early experience
Alejandro Segura B.1,2 ; Germán Gutiérrez
Universidad Nacional de Colombia, Bogotá, Colombia
RESUMEN
Se estudiaron los efectos de la experiencia alimenticia temprana en el desarrollo de conductas cooperativas en una tarea instrumental en ratas. Los sujetos fueron asignados aleatoriamente a dos grupos. En uno, los sujetos tuvieron libre acceso al alimento, condición análoga a un ambiente rico en alimento. En otro, los sujetos fueron expuestos a cantidades limitadas de alimento, representando un hábitat pobre en recursos alimenticios. En la primera fase experimental se expuso a los sujetos a una situación de elección entre respuestas cooperativas e individuales. El refuerzo obtenido por el trabajo compartido proporcionó 4 veces más de lo recibido por el trabajo individual. En la segunda fase, los sujetos obtuvieron 1 refuerzo tanto por sus respuestas individuales, como por sus respuestas compartidas. En la tercera fase, los sujetos recibieron 4 refuerzos por cada respuesta individual y 1 refuerzo por cada respuesta cooperativa. Los sujetos con experiencia temprana de escasez de alimento presentaron tasas de respuestas cooperativas más altas que los sujetos expuestos a condiciones de abundancia de alimento.
Palabras clave: Conducta social animal, Elección, Cooperación, Ratas.
ABSTRACT
We studied the effects of early experience in the development of cooperative behaviors in an instrumental task in rats. In the first experimental phase, subjects were exposed to a choice between a cooperative and a non cooperative task. By cooperating subjects obtained 4 times more food than working individually. In a second phase, all experimental subjects earned one reinforcement for each type of response. In the third phase, all experimental subjects earned four reinforcement units for responding individually and one unit for responding cooperatively. Subjects with an early experience of food deprivation presented rates of higher cooperative response than subjects early exposed to food abundance. We conclude that early experience of food abundance or deprivation influence the later development of cooperative behavior.
Keywords: Animal social behavior, Choice, Cooperation, Rats.
La investigación realizada desde el aprendizaje social, tomando como base el enfoque del análisis experimental, ha llevado a desarrollar diferentes estrategias y modelos que han permitido reproducir conductas sociales en ambientes controlados de laboratorio (Baker & Rachlin, 2002; Catania, 1992; Galef, Whiskin, & Bielavska, 1997; Gutiérrez, 1998; Pearce, 1997; Skinner, 1975). Por medio de estos estudios se han logrado observar varias conductas generadas por la interacción de dos o más organismos como comportamientos de cohesión, reciprocidad, conductas de observación, imitación, competencia y cooperación.
Desde el aprendizaje social, como lo señala Catania (1992), se pueden identificar dos formas de estudiar la interacción de los organismos. La primera, es el aprendizaje a través o alrededor de los otros. En este tipo de interacción social la conducta de un organismo puede funcionar como estímulo discriminativo para el otro organismo. La función discriminativa de la conducta de un organismo puede ser más importante que el carácter discriminativo que puede proveer un evento o objeto inanimado; por ejemplo, la inversión parental, en recursos y cuidados, que hace un sujeto al identificar correctamente a su descendencia y no confundirlos con los descendientes de otros sujetos. Así mismo, gran parte de la comunicación animal se realiza gracias a que un organismo emite una conducta (estímulo) que afecta la conducta del otro. En el ambiente natural, gran parte de la conducta es determinada por la función discriminativa de la conducta de otro organismo, como sucede con la relación entre presa y depredador; en este sentido, discriminar la propia conducta de la conducta de los demás, puede ser una ventaja evolutiva. Para los organismos sociales, el aprendizaje a través de los otros es muy importante, debido a que buena parte de la interacción y cohesión social está determinada en la correcta identificación de claves conductuales que permite generar contacto social o inclusive emerger conductas cooperativas, como lo afirma De Waal (1989, citado por Catania, 1992). La segunda forma de estudiar la conducta social, es el aprendizaje desde los otros. En esta clasificación se enfatiza la conducta que se puede aprender de otro organismo básicamente por la observación e imitación de la conducta emitida por éste (para una revisión detallada, véase Arriaga et al., en prensa). Un organismo puede elegir algún alimento específico por medio de la observación de preferencias en congéneres (ver Galef, Beck, & Whiskin, 1991; Galef & Whiskin, 1995; McQuoid & Galef, 1992). En este tipo de aprendizaje el observador, al identificar las consecuencias de la conducta emitida por un organismo, puede aprender a emitir u omitir la ejecución de dicho comportamiento.
Como se ha expuesto brevemente, el estudio de la interacción social en los organismos tiene dos vertientes principales: el aprendizaje de conductas sociales cuando la conducta del otro funciona como estímulo discriminativo y el aprendizaje de distintos comportamientos por medio de la observación e imitación de la conducta que otros, previamente han ejecutado. La presente investigación se puede ubicar en la primera distinción; es decir, dentro del aprendizaje social a través, o por medio de la interacción con el otro. La conducta de interacción cooperativa, puede emerger como señala De Waal (1989, citado por Catania, 1992) en un contexto dónde la identificación de claves comportamentales emitidas por otros sujetos les permitan asociarse en situaciones específicas. Sin embargo, un organismo puede realizar o no una asociación cooperativa; por esta razón, en esta investigación se reproduce, en un ambiente controlado de laboratorio, una situación donde los organismos pueden elegir entre cooperar o emitir conductas individuales, pero recibiendo mayor beneficio por el trabajo cooperativo que por la conducta individual; esto análogamente a las condiciones en el ambiente natural y siguiendo a De Waal y Harcourt (1992) quienes afirman que para que los organismos emitan comportamientos altruistas y cooperativos, esta conducta debe generar un beneficio mayor al recibido por conductas egoístas. Así mismo, en el presente estudio se permite a los sujetos interactuar de forma más directa al diseñar una situación donde la interacción cooperativa se realiza en un manipulando compartido, donde los sujetos pueden responder de acuerdo con las respuestas del otro y de esta manera se observa más claramente el carácter discriminativo de la conducta. Como lo ha señalado Catania (1992) la discriminación social realizada por organismos de diferentes especies tiene varias funciones como la identificación de jerarquías en la defensa del territorio y el mantenimiento de la cohesión del grupo, cuando intrusos interfieren en la distribución de fuentes limitadas de alimento; es en estos contextos, cuando la conducta cooperativa puede emerger como estrategia evolutiva permitiendo que los sujetos de esta forma defiendan los recursos. Como se observa en el anterior párrafo, al estudiar la conducta cooperativa es necesario identificar cómo la exposición a fuentes limitadas de alimento interviene en la aparición posterior de comportamientos cooperativos, por medio del presente estudio se identifica el valor que esta variable motivacional ejerce sobre este tipo de comportamiento social.
Métodos de Investigación de la Interacción Social y la Conducta Cooperativa
Son diversos los estudios interesados en la identificación de las variables que determinan la aparición de conductas de interacción social, dentro de ellos encontramos un grupo de investigaciones clásicas como la de las palomas que jugaban ping-pong y las palomas colaboradoras (Skinner, 1975) también otros estudios donde las palomas lograban comunicar al otro su estado interno (Lubinski & Thompson, 1987) o ratas que mediante la manipulación de un manipulando compartido colaboraban para obtener un refuerzo (Gutiérrez, 1998). Otro tipo de investigaciones se han centrado en el estudio de conductas de observación e imitación (Heyes & Dawson, 1990) o conductas reforzadas por estímulos sociales (Davis & Perusse, 1998; Swartz & Rosenblum, 1980). Finalmente hay otro grupo de investigaciones orientadas a la identificación de algunas variables responsables en la aparición de conductas sociales como la elección de alimento (Galef et al., 1991; Galef & Whiskin, 1995, 2001; Galef et al., 1997; Krause, Hartmann, & Pritchard, 1999; McQuoid & Galef, 1992) y la experiencia temprana (Chorpita & Barlow, 1998; González & Fleming, 2002; Matthews & Robbins, 2003; Shultz & Lore, 1993).
Como se ha expuesto anteriormente hay diversos estudios interesados por la conducta social y de la revisión detallada de estas investigaciones es posible identificar al menos, dos métodos específicos para abordar este tipo de investigaciones. El método de los sujetos observadores y los sujetos demostradores (SO-SD) (ver Figura 1), desarrollado principalmente por Bennett Galef y sus colaboradores y que caracteriza algunas de las investigaciones anteriormente citadas. El segundo método empleado es el modelo del dilema del prisionero (DP).
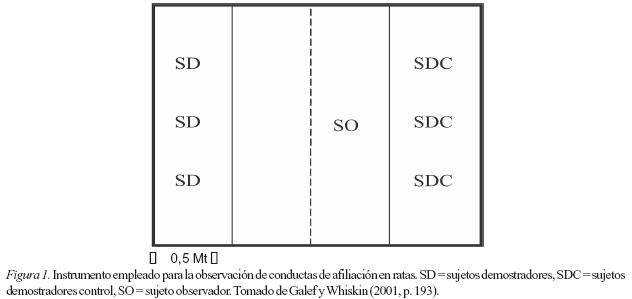
Con el método (SO-SD), se ha logrado desarrollar la investigación de uno de los campos del aprendizaje social, el aprendizaje desde los otros. Con este método se han identificado algunas de las variables que determinan que un organismo (SO) aprenda comportamientos sociales a partir de la observación de sus congéneres (SD) (ver Figura 1). Específicamente, se ha encontrado relación entre experiencias tempranas de privación alimenticia y elección de comidas especificas por medio de la observación de congéneres adultos. En síntesis, con este modelo se ha identificado la elección de los sujetos (por medio de la observación) con base en aquello que otros anteriormente han elegido.
Al revisar las investigaciones realizadas sobre conductas sociales, específicamente conductas de cooperación en los diferentes organismos, se encuentra que en las últimas décadas, los autores se han aproximado al estudio de algunas conductas sociales como competitividad, cohesión y cooperativismo adoptando como modelo el dilema del prisionero (DP) (Baker & Rachlin, 2002; Insko, Kirchner, Pinter, Efaw, & Wildschut, 2005; Komorita, Parks, & Hulbert, 1992; Wildschut, Gaertner, & Insko, 2002). Con este método se han desarrollado las investigaciones en el área del aprendizaje social a través de los otros; es decir, cuando la conducta del otro funciona como estímulo discriminativo. La interacción entre dos o más organismos puede estar determinada por los efectos y las claves discriminativas de la conducta de los sujetos que intervienen en la interacción; algunas veces, esta interacción genera un dilema. Un dilema social puede definirse como una situación en la que dos o más personas se encuentran en una relación interdependiente, en donde cada uno de los organismos entra en conflicto entre maximizar el interés personal (egoísmo) o maximizar los intereses colectivos (cooperación) (Boyd, 1992; Komorita et al., 1992). El DP puede ser explicado por medio de la teoría de juegos. Como es señalado por Keren y Raub (1993) "La teoría de juegos puede ser usada en condiciones específicas bajo las cuales los individuos racionales pueden elegir entre dos posibles actos" (p. 429). El origen de la teoría de juegos se remonta a la publicación del artículo de John von Neuman "Theory of Parlor Games" en 1928. En este artículo, Neuman presentó una primera explicación matemática (teorema minimax) de las elecciones tomadas por los "jugadores" (Poundstone, 1992). Sin embargo, es en 1944 cuando Neuman y Morgenstern presentaron una explicación teórica mas completa con "The Theory of Games and Economic Behaviour". Desde ese entonces, esta teoría se ha transformado en una herramienta útil para los científicos sociales interesados en explicar el componente racional implícito en toda interacción social. Desde la concepción de Neuman y Morgenstern (Hargreaves & Varoufakis, 1995), podemos comprender un juego como "una interacción, entre agentes, gobernada por un conjunto de reglas que especifican los posibles movimientos de cada participante y cada combinación posible de dichos movimientos" (p.1). En este sentido, un juego está comprendido por los participantes, sus decisiones y las consecuencias de sus actos; obviamente, el aspecto importante en esta concepción es que las consecuencias obtenidas no dependen sólo de las propias respuestas sino de los actos de los otros y con frecuencia de la incertidumbre producida por el desconocimiento de lo que los otros van a hacer.
En el DP (ver Figura 2) una de las elecciones lleva a recibir una consecuencia benéfica para el individuo, ignorando, lo que los demás individuos puedan recibir. Esta elección es conocida como deserción (D). La otra elección produce un efecto benéfico para ambos sujetos, en caso que ambos elijan dicha opción. Esta conducta es conocida como cooperación (C).

El DP se puede representar gráficamente en una matriz con todas las posibles combinaciones de elección realizadas por los dos sujetos. El "juego" consiste básicamente en que cuando ambos sujetos desertan (P-P) reciben la menor cantidad de refuerzo (para cada uno de los sujetos cuatro refuerzos, según se observa en la Figura 2); en contraste, cuando ambos cooperan (R-R) reciben la mayor cantidad de refuerzo (ocho refuerzos para cada uno, según se observa en la Figura 2); también se puede observar que un sujeto coopere y el otro no (S-T y T-S), en esta situación aquel que coopera recibe muy poco en relación con el que deserta, aquí obviamente cooperar es castigado (P), razón por la cual se puede observar que la conducta de los sujetos está determinada por el tipo de elección que se hace momento a momento.
Bajo estas condiciones se ha observado que las personas no suelen ser racionales al elegir sus respuestas y predominantemente emplean estrategias individuales en las distintas condiciones; sin embargo, no siempre cooperar es la respuesta más racional. Es importante aclarar que una elección es racional al evaluar sus costos y sus beneficios; es decir, de un conjunto posible de resultados la opción más racional es aquella que genera un bajo costo y a su vez representa un mayor beneficio, ya sea éste a inmediato o largo plazo. De esta manera, en una situación específica, el trabajar individualmente puede ser la estrategia racional; es decir, si cumple con el criterio de ser la opción menos costosa y más beneficiosa, asimismo en otras situaciones cooperar puede ser la opción más racional si cumple con las anteriores características. Esto puede ser más claro si observamos las diferentes formas de presentación empleadas bajo el modelo del DP:
Juegos de una Sola Presentación
Esta forma de presentar el DP se caracteriza por ser juegos de un solo ensayo o una única presentación; es decir que suele ser empleado para estudiar situaciones y dilemas sociales que suelen presentarse una sola vez en la vida. En este tipo de arreglos no hay aprendizaje por la experiencia debido a ser situaciones únicas de forma que los participantes, ya sean sujetos animales o humanos, muy probablemente no se conocen y menos aun tienen posibilidad de predecir la respuesta del compañero; en esta condición la respuesta racional es trabajar individualmente.
Juegos de Presentaciones Finitas
En esta condición los participantes son expuestos a varios ensayos o presentaciones definidas de un juego. En este tipo de arreglo los participantes aprenden con cada nuevo ensayo cómo deben responder para maximizar sus beneficios, pues constantemente están recibiendo retroalimentación de la opción elegida por su compañero. Bajo este arreglo la opción más racional es responder de acuerdo con lo que el compañero respondió en el último ensayo; es decir, si cooperó en la última presentación, también se debe cooperar. Sin embargo, al ser un número de presentaciones definidas normalmente los participantes en los últimos ensayos suelen responder de forma individual y así no arriesgarse a recibir un menor beneficio.
Juegos de Presentaciones Indefinidas
En esta forma de presentación hay una mayor impredictibilidad de la opción elegida o respuesta realizada por el compañero, pues no hay un número determinado de presentaciones o ensayos. En estas condiciones se pueden generar mayor número de respuestas cooperativas. La elección racional en juegos de presentaciones indefinidas es que ambos sujetos respondan de forma cooperativa en todos los ensayos debido a que no hay certeza sobre cual será la ultima presentación.
Como se ha expuesto hasta aquí, el interés por el estudio del comportamiento social y en particular por la cooperación entre individuos ha sido abordado mediante el uso de diversas estrategias metodológicas. En un comienzo las investigaciones se centraron en estudiar conductas de observación e imitación. Posteriormente, se ha abordado la conducta social cooperativa a partir de modelos como el dilema del prisionero y el modelo del sujeto observador y el sujeto demostrador. En estos modelos, la conducta de cada uno de los organismos se estudia utilizando manipulandos separados. Bajo estas condiciones, la conducta observada es discreta y limitada y es difícil interpretar dichas conductas como respuestas auténticamente cooperativas. Si bien investigadores del comportamiento animal han reportado numerosas instancias de cooperación en observaciones naturalistas, las condiciones de desarrollo y mantenimiento de dichas respuestas no han sido extensamente documentadas y explicadas. Algunos estudios han logrado reproducir este tipo de respuestas en el laboratorio, pero como se señaló antes, las estrategias metodológicas desarrolladas hasta ahora dejan algunas dudas sobre la validez de las medidas utilizadas. El presente estudio tenía dos objetivos fundamentales. Primero, desarrollar una estrategia metodológica válida que permitiera el estudio de respuestas cooperativas en ratas. Segundo, explorar la relación entre las experiencias tempranas de abundancia y escasez y el desarrollo de respuestas cooperativas y respuestas individuales en esta especie. Para abordar estos objetivos, se diseñó un experimento donde los sujetos podían elegir entre cooperar o trabajar individualmente, pero reproduciendo conductas continuas, análogas a las condiciones naturales. De acuerdo con varios autores, una condición necesaria para el desarrollo de conductas cooperativas es que dichas conductas provean al animal una ventaja en relación con la realización de conductas individuales. Así, el estudio se diseñó para que en una de sus fases experimentales, la conducta cooperativa generara un mayor beneficio que la conducta individual. Las otras fases experimentales del estudio fueron diseñadas como control de esta situación.
Método
Sujetos
Se utilizaron 4 ratas albinas macho (Rattus norvegicus) en buen estado de salud y de 3-4 semanas de edad al inicio del experimento. Los sujetos fueron ubicados individualmente en viviendas de acrílico de 60cm de largo, 40cm de ancho y 30cm de alto, cada vivienda tenía un nido en acrílico de 25cm de largo, 15cm de ancho y 15cm de alto, un comedero y un bebedero.
Instrumentos
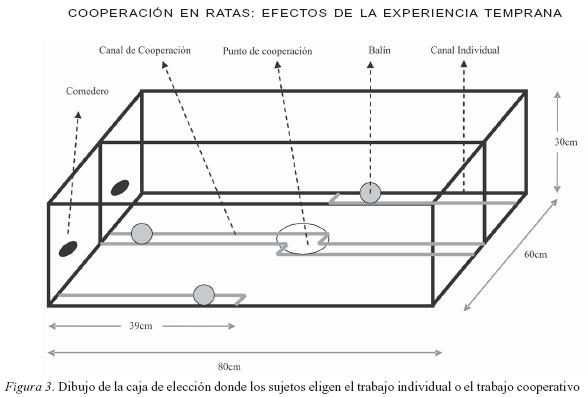
Caja experimental: Se diseñó y construyó una caja de elección de 80cm de largo, 60cm de ancho y 30cm de alto (ver Figura 3). El piso y las paredes laterales fueron hechos en madera, las paredes frontales y la división central fueron de vidrio; así, la caja fue dividida en dos compartimentos, uno para cada sujeto. En el piso se hicieron cuatro canales en aluminio de 39cm de largo y 2.4cm de ancho, dos en cada compartimiento. En los canales, un balín en acero de 2.25cm de diámetro y 55grs de peso, pudo ser deslizado al ser empujado por la rata. En la parte frontal de cada uno de los compartimientos se dispuso un canal individual. El otro canal de cada uno de los compartimientos fue dispuesto en la parte central de forma que estaba acoplado al canal del compartimiento siguiente (canal de cooperación) permitiendo que ambos sujetos realizaran las respuestas cooperativas en él. Para cada uno de los canales se instaló un dispensador de alimento donde los sujetos recibían el refuerzo alimenticio inmediatamente después de emitir las conductas (ver Figura 3). También se emplearon cronómetros, planillas de registro, videocámara y demás instrumentos necesarios para la medición y observación de la conducta.
Procedimiento
El experimento se realizó en 6 fases. En la primera fase (Habituación) se seleccionaron cuatro ratas y se distribuyeron en cuatro cajas, un sujeto por vivienda. Las condiciones de alimento sólido y líquido fueron iguales para todos los sujetos (libre acceso). Esta etapa duró dos semanas; durante este periodo y en las siguientes fases se registró el peso diario de cada una de las ratas para obtener la curva de crecimiento de los sujetos (ver Figura 4). En la segunda fase (Experiencia Temprana) se asignaron los sujetos aleatoriamente a dos grupos. Cada grupo fue compuesto por 2 sujetos. En el grupo A, abundancia temprana, los sujetos fueron expuestos inicialmente a las condiciones características de un ambiente rico en recursos; es decir, tuvieron libre acceso a la comida. En el grupo B, escasez temprana, los sujetos fueron expuestos a cantidades limitadas de alimento, es decir, el hábitat representó un ambiente pobre en recursos. Estos sujetos tuvieron una hora diaria de acceso al alimento (concentrado purina fiel) la cantidad de comida consumida durante esa hora por cada uno de ellos sirvió como medida para dejar una cantidad igual al finalizar la hora de consumo. A éste procedimiento se le denominó guardado de comida. Por ser ésta una fase de exposición temprana, los sujetos estuvieron en ésta condición durante cinco meses, de forma que su desarrollo físico coincidió con el desarrollo de esta fase. Al finalizar la segunda fase las ratas ya eran adultas. En la fase de Moldeamiento (tercera fase) se provocó el estado motivacional de privación para todos los sujetos. El índice de privación (85% de su peso corporal) fue uniforme para todos los sujetos y se mantuvo hasta el final del experimento. En esta condición todos los sujetos de ambos grupos (A y B), fueron expuestos a la caja de elección. Se esperaba que los sujetos aprendieran a desplazar el balín de un extremo al otro, en cada uno de los canales. Para lograr esta conducta se realizó un entrenamiento de aproximaciones sucesivas; esto es, que cada respuesta que se acercaba a la respuesta final deseada (llevar la esfera hasta el final del canal) fue contingentemente reforzada. El programa de reforzamiento utilizado en esta fase fue de razón fija 1 (RF1); es decir, se empleó un procedimiento de correlación positiva, relación de ejecución de la respuesta meta y la administración de alimento (Pérez- González, 1994), hasta que los sujetos respondieron consistentemente. Es importante señalar que los ensayos fueron continuos; es decir, cada vez que el sujeto llevó el balín a uno de los extremos del canal, recibió el reforzador, al haber cumplido con el anterior criterio comenzaba un nuevo ensayo. Esta fase estuvo compuesta por 4 condiciones de entrenamiento: 1) Entrenamiento en el canal individual, de la conducta de empujar el balín desde el extremo lateral al centro (para los sujetos equivale llevar el balín de derecha a izquierda), al cumplir con el criterio, el experimentador devolvía el balín al punto inicial. Por esta razón este tipo de entrenamiento fue de ensayos discretos; 2) Entrenamiento, en el canal individual, de llevar el balín del centro al extremo lateral (de izquierda a derecha); al igual que en el entrenamiento anterior los ensayos fueron discretos; 3) Entrenamiento de la conducta final deseada en el canal individual; es decir, que los sujetos ejecuten la respuesta en el canal en ambos sentidos. Al llevar el balín de un extremo al otro (operante) el sujeto recibe el refuerzo e inmediatamente comienza un nuevo ensayo sin la intervención del experimentador, el sujeto debe devolver el balín al otro extremo del canal donde también recibe refuerzo. En esta condición los ensayos son continuos; y, 4) Entrenamiento en el canal central, de llevar el balín del extremo al centro de la caja (o punto de cooperación, ver Figura 3) en este punto hay una conexión entre los dos compartimientos de la caja experimental permitiendo así el contacto entre los sujetos. Es en este punto donde el sujeto que empezó a desplazar el balín desde el extremo lateral del canal compartido, pasa ahora el balín al otro sujeto; éste lo recibe y lo lleva al extremo finalizando así un ensayo. Siendo ensayos continuos, esta última rata puede inmediatamente iniciar un nuevo ensayo al llevar ahora el balín del extremo al centro del canal y pasarlo al otro sujeto, y así sucesivamente.
En la fase experimental de Elección (cuarta fase) los dos sujetos de cada grupo fueron asignados siempre a un mismo compartimiento de la caja de elección, de esta forma se controló algún tipo de condicionamiento al contexto. En el canal central compartido se observó y registró la conducta de cooperación en ratas ya que para obtener el refuerzo el sujeto que inició el ensayo (comenzó a deslizar el balín) debió llevarlo hasta la mitad del canal y entregarlo al otro sujeto; a la vez, éste último debió recibir el balín en el centro y conducirlo hasta el final del canal. En este momento ambos sujetos recibieron contingentemente cuatro veces (cuatro unidades de comida) lo que recibían por trabajar en el canal individual. Los sujetos siempre pudieron responder libremente en ambos canales. En el canal individual cada ensayo consistía en llevar la esfera de un extremo al otro del canal, donde contingentemente el sujeto recibía un reforzador (una unidad de comida). En la quinta fase (Control 1), la cantidad de refuerzo obtenido por ensayo en el canal individual se mantiene (una unidad de comida) pero la cantidad de comida recibida por el trabajo cooperativo disminuye de cuatro unidades a una unidad de comida; es decir, en esta fase ambas elecciones le proporcionaron a los sujetos la misma cantidad de refuerzo. Esto se realizó para determinar cómo cambia la conducta de elección en razón al refuerzo obtenido; aquí la conducta cooperativa les proporciona la misma cantidad de alimento que el trabajo individual. Finalmente, en la última fase (Control 2) la elección de trabajar en el canal individual le proporcionó a los sujetos cuatro unidades de comida en vez de una unidad. En contraste, la elección del canal central proporcionó una unidad de comida. Con esta disposición se logra balancear la fase de elección de forma que se pueden observar los efectos del refuerzo en la ejecución de conductas cooperativas.
Resultados
Como se observa en la curva de crecimiento de ambos grupos (Figura 4), en la fase de Habituación se encontró que los grupos eran muy similares en su peso; inclusive, el grupo de privación temprana tenía un peso ligeramente mayor. En contraste, en la fase de Experiencia Temprana la situación cambió debido a que los grupos son expuestos a condiciones distintas (abundancia y escasez de alimento); en consecuencia, el grupo expuesto a privación (escasez) mantiene un peso 50grs inferior al grupo expuesto a saciedad (abundancia). De igual manera en la fases Experimentales (Elección, control 1 y control 2) la curva de desarrollo cambia nuevamente debido a que los sujetos de ambos grupos son expuestos a privación del 85% de su peso corporal, esto explica la caída de las pendientes de ambas curvas en los pesajes 104 a 110 (ver Figura 4), para luego estabilizarse con una pendiente cercana a cero. Nótese sin embargo que en esta fase experimental se mantiene una diferencia entre grupos de 30grs de peso aproximadamente.
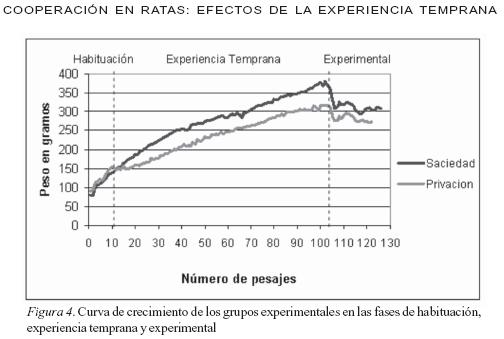
En la Figura 4 se compara el peso promedio de cada uno de los grupos, se puede observar que al inicio de la fase de experiencia temprana (pesajes 10 a 18), ambos grupos tiene un peso similar, pero la pendiente de crecimiento del grupo expuesto a privación es casi cero; en contraste, la pendiente del grupo expuesto a abundancia se mantiene igual a la fase de habituación. Desde el pesaje número 20 en adelante se puede ver que ambas curvas se separan manteniendo una diferencia entre ambos grupos de 50grs de peso aproximadamente, hasta el final de la fase de experiencia temprana (pesaje 103). Como es representado en esa gráfica el efecto de la experiencia temprana se ve reflejado en la diferencia de peso en cada uno de los grupos.
En síntesis, en la fase de experiencia temprana se encontró que aquellos organismos expuestos a condiciones similares tienen un índice de crecimiento similar, como lo visto en los sujetos que conforman un mismo grupo. Sin embargo, cuando las condiciones a las que son expuestos los organismos son diferentes (opuestas) como sucedió con la exposición de unos sujetos a ambientes ricos en recursos alimenticios y otros a ambientes pobres en recursos, el crecimiento de los sujetos de distintos grupos es diferente.
En la fase de Moldeamiento se encontró que el aprendizaje de la conducta operante en el canal individual fue muy similar en todos los sujetos y entre los dos grupos; el aprendizaje fue medido en el tiempo empleado (número de sesiones) en ejecutar la conducta correctamente. En contraste, el aprendizaje de la conducta de interacción cooperativa fue diferente para los dos grupos, debido a que el número de sesiones empleadas por los sujetos del grupo expuesto a abundancia temprana, en ejecutar correctamente la conducta final deseada, fue de 22 sesiones más que las empleadas por los sujetos expuestos a escasez temprana en aprender correctamente la respuesta. Así mismo, la tasa promedio de respuestas en cada una de las condiciones de moldeamiento en el canal individual para cada uno de los grupos fue similar. Al contrario, la tasa obtenida en el moldeamiento en el canal central fue distinta en ambos grupos. El grupo expuesto a abundancia temprana presentó un promedio de 20 respuestas más por sesión que las realizadas por el grupo expuesto a escasez temprana. En general, en esta fase de moldeamiento los sujetos del grupo expuesto a abundancia temprana presentaron una mayor activación física (movimientos rápidos y poco coordinados al desplazar el balín); en contraste, los sujetos expuestos a escasez temprana presentaron una menor activación y mayor coordinación de sus movimientos; ésta condición permitió que los sujetos de éste grupo cumplieran en un menor número de sesiones con todos los criterios del moldeamiento en cada uno de los entrenamientos (llevar el balín de un extremo al otro, en el canal individual, y llevar el balín de un extremo al centro pasándolo al otro sujeto en el canal central). En contraste, el grupo A necesitó, en toda la etapa de moldeamiento, 29 sesiones más que el grupo B.
En la fase de Elección el trabajo en el canal cooperativo proporcionó cuatro unidades de comida por respuesta y el trabajo en el canal individual una unidad de comida por respuesta. Como se puede observar en las Figuras 5 y 6 todos los sujetos distribuyeron sus respuestas en ambas elecciones; es decir, no hubo una elección exclusiva por alguna de las dos alternativas (trabajo individual o trabajo cooperativo). La elección realizada entre-grupos fue diferente; en contraste, la distribución de las respuestas intragrupo fue muy similar. Este hallazgo es muy importante porque cualquier tipo de explicación de la elección determinada por la experiencia temprana depende primero de la uniformidad en la ejecución de los sujetos que componen un grupo y segundo, de que hubiese diferencia en la distribución de la conducta entre los dos grupos. En esta fase también se encontró otro tipo de conducta que se denominó intento de cooperación; ésta respuesta consistió en el cambio que realizaron los sujetos de responder en el canal individual, para acercarse al canal central aunque no hubiese el balín o, si hubiese balín, no completaban la operante y cambiaban de nuevo a responder en el canal individual. También se puede interpretar esta conducta como una invitación a cooperar. Al comparar la fase de elección con la fase de Control 1 se observa un cambio en la distribución de las respuestas de todos los sujetos (ver Figuras 5 y 6). Esto sucedió debido a que la cantidad de refuerzo obtenido en ambas elecciones fue igual (1 refuerzo por respuesta). Sin embargo, se hallaron diferencias de ejecución en cada uno de los grupos debido a la experiencia temprana. Los sujetos del grupo B deprivado presentaron un aumento gradual de las respuestas en la opción individual y una disminución, también gradual, de las respuestas cooperativas. En contraste, los sujetos del grupo A saciado presentaron una distribución de las respuestas similar al observado en la fase anterior; es decir, no se hallaron cambios significativos a lo largo de las sesiones. Sin embargo, se halló un ligero aumento de las respuestas individuales y una disminución de las respuestas cooperativas. Finalmente, en la fase de control 2 el trabajo en el canal compartido proporcionó una unidad de comida por respuesta y el trabajo en el canal individual cuatro unidades de comida por respuesta. En esta fase no se encontraron diferencias en la ejecución de los sujetos de ninguno de los grupos. Esta homogeneidad en la distribución de sus respuestas se debe a que la opción de trabajo cooperativo les ofrece a los organismos una menor cantidad de alimento que el trabajo individual.
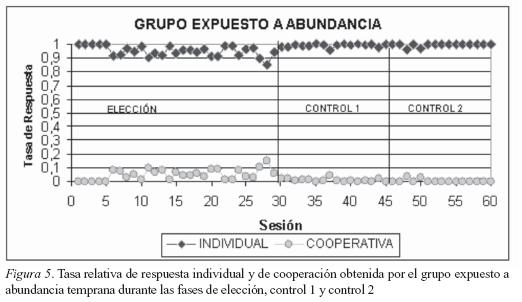
En la Figura 5 se observa que en las primeras 5 sesiones los sujetos del grupo saciado realizaron una elección exclusiva por el trabajo en la opción individual. También se puede ver que de la sesión 6 a la 30 (que hacen parte de la fase de elección) los sujetos mantienen una tasa de respuesta estable entre 0.9 para la opción individual y 0.1 para la opción de cooperación; claramente se observa una marcada preferencia por el trabajo individual sobre el de cooperación. Esta preferencia se marca aun más en las sesiones 31 a 45 (fase de control 1) y 46 a 60 (fase de control 2) donde la tasa de respuesta individual es muy cercana a 1 y la tasa de respuesta cooperativa en muchas sesiones es 0.
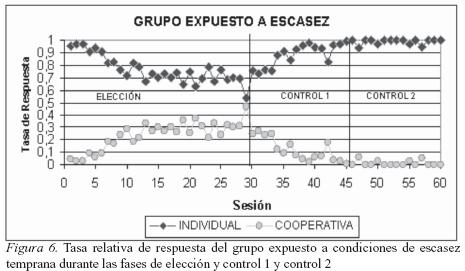
Como se puede observar en la Figura 6 los sujetos del grupo expuesto a escasez distribuyeron sus respuestas en ambas opciones (trabajo individual y trabajo cooperativo). En la fase de elección la tasa de respuesta cooperativa está entre 0.3 y 0.4 y la tasa de respuesta individual entre 0.6 y 0.7. Esto es equivalente a una distribución de 30% de trabajo cooperativo y un 70% de trabajo individual. En la fase de control 1 se observa un decremento temporal (sesión a sesión) de las respuestas cooperativas y una preferencia gradual por la opción individual. Finalmente en la última fase (sesiones 46 a 60) hay una preferencia aun mayor por el trabajo de tipo individual, ejecución muy similar a la observada en el grupo expuesto a saciedad temprana.
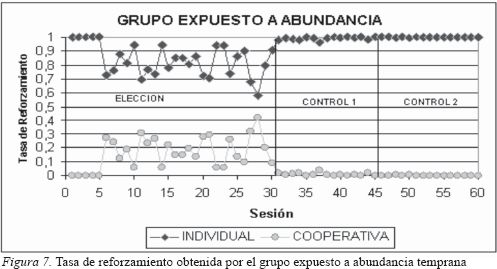
La estrategia comportamental (distribución de respuestas) del grupo saciado en la fase de elección les permitió obtener por cooperar, el 16% y por trabajar individualmente, el 84% del total de refuerzo obtenido por sus respuestas en esta fase. Es decir que, aunque la mayor cantidad de alimento por respuesta la proporcionaba la opción de cooperar (cuatro unidades), los sujetos mostraron una preferencia por responder en la opción individual que proporcionaba una unidad. En esta fase la mayor parte del alimento consumido por los sujetos fue obtenido con una estrategia de elección de trabajo individual, sin importar que el costo fuera una tasa de respuesta muy alta en dicha opción. Esta distribución se polariza aun más en las fases de control donde el 99% del refuerzo fue obtenido por el trabajo en la opción individual. Sin embargo, la distribución encontrada en estas dos ultimas fases (control 1 y control 2) es normal debido a que la cantidad de alimento por respuesta es igual o mayor en la opción de respuesta individual que en la opción de respuesta cooperativa, donde la inversión de tiempo es mayor; es esperable entonces que los sujetos tiendan a responder casi exclusivamente en la opción de menor costo.
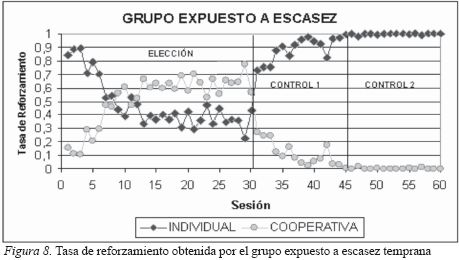
Como se puede observar en la Figura 8 los sujetos del grupo expuesto a escasez temprana en las primeras sesiones de la fase de elección obtuvieron una mayor cantidad de alimento por sus respuestas en la opción de trabajo individual que por sus respuestas en la opción de trabajo cooperativo. Sin embargo, esta distribución se invierte a partir de la sesión 9 donde el 57% del alimento que obtienen lo reciben por la distribución de sus respuestas en la opción compartida; de la sesión 13 a la 30, la tasa de refuerzo para la opción de cooperación se estabiliza por encima del 60% del total de refuerzo obtenido en esta fase. La estrategia emitida por los sujetos de este grupo expuesto a escasez temprana es contraria a la observada en el grupo expuesto a abundancia temprana, al observarse una marcada preferencia por cooperar al obtener mayor cantidad de alimento por un número reducido de respuestas, recordemos que en esta fase una respuesta en la opción de cooperación representa cuatro unidades de comida y una respuesta en la opción individual representa 1 unidad de comida. En contraste, en las sesiones 31 a 45 (fase de control 1) de nuevo la tasa de reforzamiento se invierte y la mayor cantidad de alimento es obtenido por el trabajo individual; esta distribución es esperable debido a que la cantidad de refuerzo obtenido es igual para ambas opciones (una unidad de comida por respuesta en cada opción). Sin embargo, la cantidad de refuerzo obtenido por cooperar en esta fase es del 12% (mucho mayor al observado en el grupo expuesto a abundancia). Finalmente, en la fase de control 2 la cantidad de refuerzo obtenido se estabiliza y el 99% del alimento consumido es conseguido por el trabajo individual. En esta fase el trabajo individual proporciona cuatro unidades de comida en contraste con una unidad de comida recibida por cooperar. En esta fase de control 2, la tasa de reforzamiento observada en el grupo expuesto a escasez es muy similar a la tasa de reforzamiento del grupo expuesto a abundancia (ver Figuras 7 y 8).
Discusión
El interés primordial de este estudio fue crear una estrategia o táctica metodológica que permitiera observar la existencia de un fenómeno conductual y las condiciones bajo las cuales éste fenómeno se da. En este sentido, con este experimento se logró establecer una nueva técnica que permite estudiar comportamiento social cooperativo y, al mismo tiempo, observar bajo qué condiciones los organismos cooperan. Siendo éste el objetivo central del experimento se eligieron ratas como sujetos experimentales, por ser una especie gregaria y de fácil manejo experimental. Nuestro fin no es el de generalizar los datos a ratas y humanos, en este sentido compartimos la postura, que frente a la analogía de los experimentos, propone Sidman (1973) "Nuestro problema no es el de obtener análogos, sino el de llegar a comprender tanto las ratas como a los hombres, y ser capaces de reconocer paralelismos en los procesos conductuales" (p. 37). De esta manera con este experimento se identificó un proceso conductual (comportamiento cooperativo en ratas) como punto inicial en la búsqueda de las condiciones bajo las cuales diversos organismos pueden llegar a cooperar. Asimismo, en este experimento se empleó un número reducido de sujetos porque, como se señalo antes, nuestro objetivo era reconocer un fenómeno conductual y reproducirlo en ambiente controlado. En contraste, algunos de los experimentos que emplean los modelos del dilema del prisionero y los sujetos demostradores y observadores, utilizan un número mayor de participantes porque ya se han realizado un número considerable de réplicas y en estos estudios no se busca reproducir un fenómeno conductual per-se sino realizar una generalización estadística de los datos y observar nuevas variables. Es importante aclarar que si bien el uso de herramientas estadísticas en la explicación de los datos es de gran valor, asimismo la observación y el análisis de sujetos individuales continúa siendo una estrategia metodológica válida en la formulación y comprobación de hipótesis.
En relación con los datos observados se pueden identificar tres hallazgos básicos de esta investigación. Primero, con este trabajo fue posible diseñar un modelo diferente para observar interacción social en ratas desde el aprendizaje social y específicamente el aprendizaje a través de los otros. A diferencia del modelo del dilema del prisionero donde los organismos que interactúan responden en manipulandos separados, con este estudio se logró reproducir conducta cooperativa, cuando dos organismos trabajan simultáneamente para cumplir con una respuesta.
Los hallazgos de esta investigación, al igual que las investigaciones basadas en el dilema del prisionero (DP) (revisar Baker & Rachlin, 2002; Insko, Kirchner, Pinter, Efaw, & Wildschut, 2005; Komorita et al., 1992; Wildschut, Gaertner, & Insko, 2002) permiten afirmar que los organismos tienden a distribuir sus respuestas entre el trabajo individual y el cooperativo cuando el refuerzo obtenido por cooperar es mayor que lo recibido por la elección individual; sin embargo, con este diseño, a diferencia del modelo del dilema del prisionero, se observaron conductas cooperativas cuando el beneficio por cooperar era igual a lo obtenido por trabajar individualmente (de acuerdo con lo observado en la fase de control 1en el grupo expuesto a escasez) esto sucedió porque la experiencia temprana alimenticia que viven los sujetos determina en gran medida la aparición de comportamientos cooperativos en etapas posteriores en el desarrollo.
El diseño que se empleó en esta investigación permitió, al igual que en las investigaciones basadas en el modelo de los sujetos observadores y demostradores (SO-SD) (Galef et al., 1991, 1995, 1997, 2001; Krause, Hartmann, & Pritchard, 1999; McQuoid & Galef, 1992), observar la relación entre variables alimenticias y la ejecución de conductas sociales. Sin embargo, los trabajos bajo el modelo SO-SD, se ubican dentro de las investigaciones del aprendizaje social desde los otros, específicamente el aprendizaje a través de la observación e imitación. A partir de allí se puede afirmar que los organismos aprenden a seleccionar alimentos específicos y elegir de acuerdo con la observación de las elecciones que realizan sus congéneres. En contraste, con esta investigación se logró observar que los organismos eligen cooperar de acuerdo con la retroalimentación obtenida de la interacción con sus congéneres y de la discriminación de la conducta del otro (invitación a cooperar). Es decir, con lo observado en este experimento se puede afirmar que, al igual que los trabajos basados en el modelo SO-SD, la experiencia temprana de privación de alimento determina la aparición de conductas sociales en ratas pero no solamente cuando el aprendizaje es por observación sino cuando se aprende a través de la interacción con los otros.
Segundo, con el diseño empleado en este experimento se obtuvieron conductas cooperativas en un ambiente de laboratorio y en condiciones en que los sujetos podían elegir entre cooperar o trabajar individualmente. Esta condición de elección permitió observar la distribución del comportamiento en un lapso de tiempo determinado. Además, al haberse empleado ensayos continuos se permitió que dicha distribución de la conducta fuese lo más análoga posible a las condiciones naturales donde la conducta sucede, y especialmente situaciones donde emergen comportamientos cooperativos.
Tercero, se logró identificar la relación entre un estado motivacional como la experiencia temprana de privación o escasez de alimento y la posterior elección de conducta de cooperación en ratas. Así, aquellos sujetos que en una etapa temprana en el desarrollo vivieron situaciones precarias de alimento tienden a presentar una proporción mayor de conductas sociales de cooperación que organismos que se desarrollaron sin limitaciones de alimento.
La diferencia observada en el aprendizaje de los sujetos en la fase de moldeamiento permite sugerir que el aprendizaje de los sujetos puede estar determinado en parte, por las experiencias anteriores que han vivido; así, aquellos organismos que habitan ambientes adversos - con acceso limitado al alimento- en etapas tempranas de su desarrollo es más probable que aprendan más rápidamente tareas operantes que organismos que han habitado ambientes ricos en recursos alimenticios. Esto puede suceder, porque la experiencia temprana genera cambios conductuales como impulsividad y mayor activación en organismos expuestos a abundancia que en organismos expuestos a escasez. Así mismo, otra posibilidad es que estos organismos expuestos a escasez, por habitar en ambientes donde deben estar atentos a claves (estímulos) específicas que predicen el acceso al alimento, pueden desarrollar una mayor atención a otro tipo de estímulos en etapas posteriores de su vida y en consecuencia tener una mayor facilidad para aprender tareas operantes.
Como se observó en la fase de elección, la distribución de las respuestas que realizan los sujetos está determinada por las experiencias anteriores, en este caso, por la experiencia temprana a ambientes diferentes. Aunque se identificó una distribución diferente de la conducta entre los grupos, los sujetos de ambos grupos presentaron mayor tasa de conductas individuales sobre las conductas cooperativas. Esto sucedió porque el tiempo empleado en la ejecución de cada una de las respuestas fue diferente. El tiempo invertido por un sujeto en el canal individual fue considerablemente menor al tiempo empleado en el canal de cooperación; así mismo, los sujetos gastaron menos tiempo en consumir una unidad de comida (lo obtenido en el canal individual) que cuatro unidades de comida (obtenidos por cooperar). Estas diferencias de tiempo determinaron que los organismos cambiaran de opción, momento a momento, y tuvieran una mayor tasa de respuestas en la opción individual. En contraste, la tasa de reforzamiento en la opción de cooperar del grupo expuesto a escasez alimenticia en la fase de elección fue considerablemente mayor que la observada por trabajar individualmente. Asimismo, el haber observado diferencias en la ejecución entre grupos permite afirmar que la experiencia temprana determinó la elección de los sujetos.
Como también se observó, cuando el beneficio obtenido por el trabajo individual y el trabajo grupal es el mismo (fase de control 1), los organismos tienden a presentar mayores conductas individuales y a disminuir sus respuestas cooperativas. Sin embargo, se siguen observando diferencias en la distribución de respuestas entre los grupos (escasezabundancia) razón por la cual se puede afirmar que la experiencia temprana tiene efectos en la elección de conductas cooperativas, inclusive cuando no hay un beneficio mayor por realizar esta conducta. Así, el grupo expuesto a escasez temprana presentó un mayor promedio de respuestas cooperativas que el grupo expuesto a abundancia. Cuando el beneficio obtenido por el trabajo individual es considerablemente mayor al obtenido por el trabajo grupal (fase de control 2), la experiencia temprana ya no tiene un efecto muy claro sobre la elección y los organismos tienden a trabajar individualmente. En este caso, la conducta cooperativa se extingue y los organismos eligen la opción que mayor beneficio les proporciona. Esto es esperable, por cuanto no sería una conducta adaptativa el trabajar cooperativamente cuando se puede acceder a mayor comida trabajando individualmente e invirtiendo menor tiempo.
Finalmente, es importante resaltar que la observación de comportamiento cooperativo en ratas en ambiente de laboratorio y empleando la técnica propuesta en este estudio es un punto de partida en la investigación de este fenómeno conductual. Sabemos que la identificación de los procesos conductuales comunes que subyacen a la cooperación es un área de investigación aun por explorar; la siguiente tarea tal vez sea evaluar la fiabilidad y generalidad de los hallazgos de este estudio mediante la replicación sistemática de este experimento, utilizando los datos obtenidos y realizando nuevos experimentos que permitan obtener datos relacionados adicionales.
Referencias
Arriaga, P., Ortega, M., Meza, G., Huichán, F., Maldonado, E., Rodríguez, A., & Cruz, S. (en prensa). Análisis de los conceptos relacionados con el aprendizaje observacional y la imitación. Revista Latinoamericana de Psicología. [ Links ]
Baker, F., & Rachlin, H. (2002). Teaching and learning in a probabilistic Prisoners Dilemma. Behavioural Processes, 57, 211-226. [ Links ]
Boyd, R. (1992). The evolution of reciprocity when conditions vary. In A. Harcourt & F. De Waal (Eds.), Coalitions and alliances in humans and others animals (pp.473-492). New York, USA: Oxford University Press. [ Links ]
Catania, A. C. (1992). Learning. Englewood: Prentice Hall. [ Links ]
Chorpita, B. F. & Barlow, D. H. (1998). The development of anxiety: The role of control in the early environment. Psychological Bulletin, 124, 3-21. [ Links ]
Davis, H., & Perusse, R. (1988). Human based social interaction can reward a rats behavior. Animal Learning & Behavior, 16, 89- 92. [ Links ]
De Waal, F .B., & Harcourt, A. H. (1992). Coalitions and alliances: A history of ethological research. In A. Harcourt & F. De Waal (Eds.), Coalitions and alliances in humans and others animals (pp.1-19). New York, USA: Oxford University Press. [ Links ]
Galef, B. G., Beck, M., & Whiskin, E. E. (1991). Protein deficiency magnifies social influence on the food choices of Norway rats (Rattus norvegicus). Journal of Comparative Psychology, 105, 55-59. [ Links ]
Galef, B. G., & Whiskin, E. E. (1995). Learning socially to eat more of one food than another. Journal of Comparative Psychology, 109, 99-101. [ Links ]
Galef, B. G., & Whiskin, E. E. (2001). Effects of Caloric, Protein, and Sodium Deprivation on the Affiliative Behavior of Norway Rats (Rattus norvegicus). Journal of Comparative Psychology, 115, 192-195. [ Links ]
Galef, B. G., Whiskin, E. E., & Bielavska, E. (1997). Interaction with demonstrator rats changes observer rats affective responses to flavors. Journal of Comparative Psychology, 111, 393-398. [ Links ]
González, A., & Fleming, A. S. (2002). Artificial rearing causes changes in maternal behavior and C-fos expression in juvenile female rats. Behavioral Neuroscience, 116, 999-1013. [ Links ]
Gutiérrez, L. F. (1998). Efecto de 4 programas simples de reforzamiento sobre las respuestas de interacción grupal de 2 ratas albinas que comparten un manipulando. Tesis de Grado inédita, Universidad Nacional de Colombia, 4.5/G – 984e. [ Links ]
Hargreaves, S. P., & Varoufakis, Y. (1995). Game theory. A critical introduction. Londres: Routledge. [ Links ]
Heyes, C. M., & Dawson, G. R. (1990) A demonstration of observational learning using a bidirectional control. Quarterly Journal of Experimental Psychology, 42B, 59-71. [ Links ]
Insko, C. A., Kirchner, J., Pinter, B., Efaw, A., & Wildschut, T. (2005). Interindividual-intergroup discontinuity as a function of trust and categorization: The paradox of expected cooperation. Journal of Personality and Social Psychology, 88, 365-385.
Keren, G., & Raub, W. (1993). Resolving social conflicts through hostage posting: Theoretical and empirical considerations. Journal of Experimental Psychology: General, 122, 429-448.
Komorita, S. S., Parks, C. D., & Hulbert, L. G. (1992). Reciprocity and the induction of cooperation in social dilemmas. Journal of Personality and Social Psychology, 62, 607-617.
Krause, J., Hartmann, N., & Pritchard, V. (1999). The influence of nutritional state on shoal choice in zebrafish, Danio rerio. Animal Behaviour, 57, 771-775.
Lubinski, D., & Thompson, T. (1987). An animal model of the interpersonal communication of interoceptive (private) states. Journal of the Experimental Analysis of Behavior, 48, 1-15.
Matthews, K., & Robbins, T. W. (2003). Early experience as a determinant of adult behavioural responses to reward: The effects of repeated maternal separation in the rat. Neuroscience and Biobehavioral Reviews, 27, 45-55.
McQuoid, L. M., & Galef, B. G. (1992). Social influences on feeding site selection by Burmese fowl (Gallus gallus). Journal of Comparative Psychology, 106, 137-141.
Pearce, J. M. (1997). Animal learning and cognition: An introduction. East Sussex: Psychology Press.
Pérez-González, A. (1994). Psicología del aprendizaje: Manual de laboratorio. Bogotá: Fondo Nacional Universitario.
Poundstone, W. (1992). Prisoners Dilemma. New York, USA: Doubleday Press.
Shultz, L. A., & Lore, R. K. (1993). Communal reproductive success in rats (Rattus norvegicus): Effects of group composition and prior social experience. Journal of Comparative Psychology, 107, 216-222.
Sidman, M. (1973). Tácticas de investigación científica. Barcelona: Fontanella.
Skinner, B. F. (1975). Registro acumulativo. Barcelona: Fontanella.
Swartz, K. B., & Rosenblum, L. A. (1980). Operant responding by Bonnet Macaques for Color Videotape Recordings of Social Stimuli. Animal Learning and Behavior, 8, 311-321.
Wildschut, T., Gaertner, L., & Insko, C. A. (2002). Intragroup social influence and intergroup competition. Journal of Personality and Social Psychology, 82, 975-992.
Received 18/07/2005
Accepted 30/05/2006
1 Dirección: Apartado 76879, Bogotá, Colombia. E-mail: alejandrosegurab@yahoo.es; ggutierrez@rlpsi.org
2 Prémio Estudantil SIP 2005
